
The requested URL /topic.htm was not found on this server.
Учение Магнуса о статических рефлексах знаменует собой новейшие успехи экспериментальной физиологии лабиринта и стволовой части головного мозга и представляет одну из важнейших и интереснейших проблем современной ото - неврологии.
Изложение учения Магнуса вообще, тем более - краткая его интерпретация, представляет довольно трудную задачу. В целях наибольшей наглядности и ясности изложения мы будем следовать в дальнейшем ходу самих экспериментов Магнуса, ограничиваясь преимущественно только выводами, по возможности опуская частности и детали.
Все статические рефлексы Магнуса разделяются на две самостоятельные трупы: рефлексы положения (Haltungsreflexe) и установочные рефлексы (Stellreflexe).
Отправным пунктом экспериментов Магнуса послужило животное, находящееся в состоянии "децеребрированной ригидности" Шеррингтона. Если у животного перерезать мозговой ствол между продолговатым мозгом и четверохолмием, то получится следующее: все конечности животного, а также мышцы шеи и туловища вытянутся в сильнейшей экстензии; такое животное, если его поставить, будет стоять совершенно неподвижно, оно не способно удерживать равновесие и производить какие-либо движения. Это и есть состояние "децеребрированной ригидности". Описанное явление есть состояние рефлекторное - оно исходит от проприоцептивных раздражений чувствительных нервных окончаний самих мышц, что и служит началом рефлекса; если перерезать задние, т.е. чувствительные корешки - ригидность исчезает.
Если такому животному повернуть голову, то конечности также изменят свое положение. Эта связь между положением головы и положением конечностей имеет определенную закономерность и зависит от двух условий: как меняется положение головы вообще в пространстве и - по отношению к туловищу. Для того, чтобы изучить влияние абсолютных изменений положения головы на положение конечностей, голову животного фиксируют к туловищу помощью гипсовой повязки. Если теперь вращать животное вокруг, напр., битемпоральной оси, то окажется, что максимум тонического экстензорного состояния конечностей будет соответствовать положению животного на спине, минимум - обратному положению (на 180°).
Это есть рефлекс, исходящий из лабиринта - тонический лабиринтный рефлекс на конечности (рис. 18); если разрушить лабиринты - явление исчезает, т.е. вращение животного теперь не изменит положения конечностей.
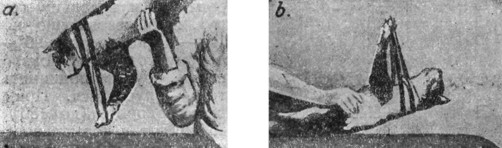 |
Рис. 18. Тонический лабиринтный рефлекс на конечности (по Magnas'y). |
Дальнейшие опыты с центрифугированием животного по способу Виттмаака показали, что этот лабиринтный рефлекс исходит от слуховых пятен - разрушение их уничтожает рефлекс, а с помощью специальных анатомических исследований была установлена связь его именно с отолитовым аппаратом utriculus'a. Оба отолита последнего (у кошки) лежат в одной горизонтальной плоскости как при максимальном, так и при минимальном сокращении мышц-экстензоров, и переход одного в другое зависит от поворота животного на 180°. Отолит (кристаллы углекислой извести) располагается, как известно, на волосках клеток пятен. При положении головы, соответствующем raaximum'y тонического рефлекса, отолит висит на волосках клеток (оттягивает их), при обратном, на 180° - отолит лежит (давит) на них, что соответствует minimum'y рефлекса. Из этого следует, что характер нагрузки отолитом волосков клеток maculae utriculi и есть основа тонического лабиринтного рефлекса на конечности. Важно, что разрушение одного лабиринта в этом опыте не уничтожает рефлекса; следовательно, отолитовый аппарат utriculus'a каждой стороны имеет отношение ко всем четырем конечностям, разрушение же обоих лабиринтов прекращает эту реакцию. Если теперь у животного с удаленными лабиринтами снять гипсовую повязку с тем, чтобы можно было вращать голову по отношению к туловищу, то окажется, что тому или иному положению головы будет также соответствовать определенное положение конечностей. При сгибании головы передние конечности будут сгибаться, а задние - разгибаться; наоборот - при разгибании головы передние конечности вытянутся, а задние сократятся. Если поворачивать голову животного в стороны, то те конечности, куда повернута голова, будут разгибаться, а . конечности другой стороны, наоборот, сгибаться. Это есть тонические шейные рефлексы на конечности. В основе их лежат проприоцептивные раздражения нервных окончаний шейных мышц; перерезка шейных чувствительных корешков устраняет рефлексы.
Если же у животного лабиринты не разрушены, то положение конечностей, соответствующее тому или иному положению головы, будет зависеть теперь от суммирования двух конкурирующих влияний лабиринтов и шейных мышц. Указанные отношения в значительной степени усложняются тем обстоятельством, что лабиринты, в свою очередь, влияют также и на шейные мышцы; влияние это односторонне и гомолатерально; при разрушении только одного лабиринта голова повернута в сторону другого (здорового), вследствие преобладания тонуса соответствующих мышц. Степень этого тонического сокращения шейных мышц также зависит от изменения положения головы в пространстве, причем максимум и минимум этого состояния вполне совпадают с таковыми для лабиринтных рефлексов на конечности. Следовательно, описанные тонические лабиринтные рефлексы на мышцы шеи стоят в той же самой зависимости от отношения отолитов к волоскам клеток утрикула.
Таким образом, воздействие лабиринтов одновременно осуществляется и на конечностях (обеих сторон) и на мышцах шеи (своей стороны). А так как шейные мышцы, как мы видели, определенным образом влияют на положение конечностей, то, следовательно, лабиринт может оказывать воздействие на мышцы конечностей также и посредственным - через них - путем.
|
The requested URL /down.htm was not found on this server.