
The requested URL /topic.htm was not found on this server.
Преддверный корешок слухового нерва, вступая в Варолиев мост, глубоко проникает в вещество мозга кнутри от веревчатого тела в направлении ко дну IV желудочка (рис. 6). Достигнув внутреннего отдела нижней мозжечковой ножки, преддверный корешок отдает восходящие и нисходящие волокна. Нисходящая часть корешка спускается в составе мозжечковой ножки, достигая спинного мозга. Большинство его волокон оканчивается в клетках так наз. треугольного или заднего вестибулярного ядра (n. vestibularis triangularis), лежащего на дне ромбовидной ямки и соответствующего здесь области trigonum acustici (см. рис. 2, area acustica). Между волокнами самого корешка различается, кроме того, ядро нисходящего корешка (Роллера), а между отдельными пучками мозжечковой ножки располагается ядро Дейтерса с очень крупными клетками. Здесь также оканчивается часть волокон преддверного нерва.
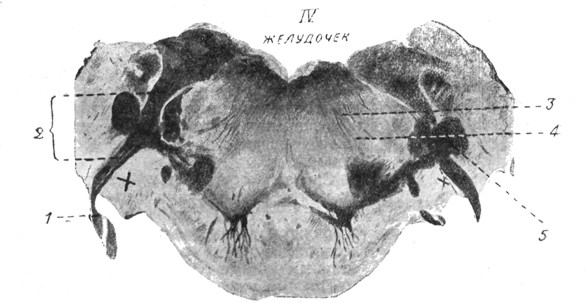 |
Рис. 6. Вход преддверного корешка слухового нерва в продолговатый мозг на границе с Варолиевым мостом (по Marburg'y). |
Волокна восходящего корешка преддверного нерва оканчиваются, главным образом, в ядре Бехтерева, состоящем из более мелких клеток и лежащем у наружного угла дна четвертого желудочка, и поэтому также называемом nucl. angularis.
Кроме того, часть волокон преддверного нерва, не прерываясь в указанных ядрах, направляется прямо в мозжечок, где и оканчивается в одном из его ядер (кровельном ядре, быть-может, также и в других; см. о мозжечке).
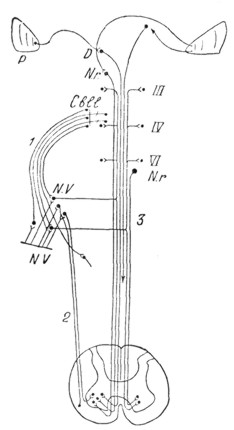 |
Рис. 7. Связи вестибулярных ядер с мозжечком (1), спинным мозгом (2) и ядрами глазных мышц (3). |
Существуют, невидимому, также корешковые волокна vestibularis, идущие непосредственно к ядрам продолговатого и спинного мозга. Этим, между прочим, может быть объяснено исчезновение некоторых вестибулярных рефлексов только после разрушения самого корешка преддверного нерва (Валленберг). Особого интереса заслуживают данные (Винклер), позволяющие считать, что часть волокон из преддверного корешка направляется к ядрам улиткового корешка слухового нерва и, наоборот, некоторые волокна этого последнего достигают вестибулярных ядер,- мнение, позволяющее отрицать принципиальную разницу в способе окончаний кохлеарного и вестибулярного корешков слухового нерва.(О частном значении этого факта мы еще скажем ниже в связи с некоторыми общими выводами).
Связи конечных вестибулярных ядер с другими отделами центр, нервной системы устанавливаются в трех основных направлениях: с мозжечком, выше лежащими отделами мозгового ствола и спинным мозгом (см. схему на рис. 7).
Помимо уже упомянутых прямых волокон преддверного нерва, идущих из корешка к ядрам мозжечка, все вестибулярные ядра, кроме, быть-может, Дейтерсова, посылают сюда свои отростки - этим устанавливаются тесные отношения n. vestibularis с мозжечком. Связь вестибулярных ядер со спинным мозгом устанавливается пучком Лёвенталя (f. vestibule-spinalis), берущим свое начало от больших клеток Дейтерсова ядра; располагаясь в продолговатом мозгу между оливой и n. ambiguus (ядро блуждающего нерва), он спускается в спинной мозг, где занимает место по периферии переднего столба и оканчивается преимущественно в шейной части мозга. Здесь он носит также название, по занимаемому им положению, передне-краевого пучка Лёвенталя. Некоторые допускают прямую связь этого пучка с мозжечком, что остается, впрочем, сомнительным. Но наличие связей мозжечка с ядрами преддверного нерва (см. о мозжечке), в частности - ядром Дейтерса (f.f. cerebello-vestibulares) ставит вестибуло-спинальный пучок в зависимость и от этого органа; в этом смысле он может быть отнесен к мозжечковым (экстрапирамидным) системам.
Благодаря наличию этой системы волокон осуществляется важное тонизирующее влияние преддверного нерва resр. вестибулярного аппарата на шейные мышцы, а также туловища и конечностей; этим путем осуществляются, в частности, специальные сложные рефлексы, исходящие из лабиринта (отолитов мешечков преддверия, см. рефлексы Магнуса).
Кроме этой важной системы, ядро Дейтерса посылает небольшое количество волокон в задний продольный пучок (fasc. longitudinalis dorsalis) в нисходящем направлении (чем устанавливается связь с ядром блуждающего нерва), также спускающихся в спинной мозг. Восходящие же волокна, идущие к ядрам глазных мышц, указанный пучок получает из клеток ядра Бехтерева;1 эта система (f. f. vestibule-oculomotoriae) приобретает здесь особенно важное значение, так как именно этим путем осуществляется влияние преддверного нерва на мышцы глаза: 1) нистагм при искусственном раздражении преддверного нерва (экспериментальный нистагм при вращательной, калорической и гальванической пробах) и патологических состояниях - спонтанный нистагм, и 2) компенсаторные движения глаз, не изменяющие направления взгляда при переменах положения головы (см. рефлексы Магнуса). Нистагм глазных яблок имеет исключительно важное значение в патологии внутреннего уха и соответствующих отделов центральной нервной системы. Ирритативный процесс вызывает нистагм в сторону очага, деструктивный - в здоровую сторону; симптом сопровождается реактивными движениями туловища и конечностей (падение тела и отклонение конечностей).
Как видно из предыдущего, нистагм не может быть объяснен участием ядра Дейтерса, так как последнее не стоит в связи с ядрами глазных мышц; в то же время отдельные анатомо-клинические наблюдения свидетельствуют как-будто в пользу этой возможности (Марбург). Вообще, в учении о нистагме в анатомо-физиологическом отношении остается много неразрешенных вопросов. Это, впрочем, относится в известной степени и ко всей вообще системе преддверного нерва: некоторые выводы экспериментальной физиологии, как и многие клинические факты, не могут быть объяснены наличием известных нам систем vestibularis. Как пример, можно было бы указать следующий факт из области новейшей физиологии: центром лабиринтных установочных рефлексов Магнуса является красное ядро в четверохолмии, тем не менее афферентных вестибулярных путей к этому образованию мы не знаем. (Учение Магнуса о статических рефлексах, как и их анатомические основания, будут рассмотрены нами отдельно - см. соответствующую главу).
Совершенно неизвестными также остаются для нас и центральные пути преддверного нерва и корковые центры vestibularls. Исходя из функциональных особенностей вестибулярного аппарата как органа статики и кинетики тела, многие склонны вообще отрицать существование этих центров. Но едва ли даже существует для этого действительная необходимость: решение вопроса следует, очевидно, искать в ином направлении.
Отсутствие известных нам центральных путей vestibularis вместе с топографической близостью улитки и преддверия - органов слуха и статики - дает основание интересной концепции, позволяющей с филогенетической точки зрения объединить в биологическом отношении кохлеарную и вестибулярную части нерва. Существует целый ряд фактов, которые, несмотря на кажущееся принципиальное различие между улиткой и преддверием, позволяют провести между ними известную аналогию и установить более близкую связь. Так, например, есть основание допускать, что низшие животные, не имеющие улитки, органа филогенетически более нового, или имеющие только зачаток ее (lagenula), но зато хорошо развитые полукружные каналы, способны реагировать на звуковые раздражения. У них же, несмотря на отсутствие cochleae, имеется система волокон, аналогичная боковой петле (lemniscus lat.), являющейся, как нам известно, центральным слуховым путем высших животных. Но даже у этих последних имеется соединительная ветвь внутри лабиринта, входящая в состав ramus cochlearis из дистального (вестибулярного) корешка (см. иннервацию лабиринта); некоторое количество вестибулярных волокон проникает в слуховые ядра продолговатого мозга и, наоборот - часть слуховых волокон достигает вестибулярных ядер; наконец, тождественность в строении чувствительного эпителия периферических воспринимающих органов, улитки и преддверия дополняет эту связь. Но как понять различие в способе окончаний вестибулярной и кохлеарной частей нерва у высших животных?
По существующему в настоящее время воззрению, всю вообще чувствительную способность высших животных можно разделить на два вида: корковую, neoaesthesi'ю, и подкорковую, paleoaesthesi'ю, или эпикритическую и протопатическую чувствительность (Head). С этой точки зрения вестибулярную функцию можно рассматривать, как подкорковую, т.е. paleoaesthesi'ю, а слуховую - как корковую, т.е. neoaesthesi'ю. Следовательно, в этом понимании кохлеарная и вестибулярная функции не противопоставляются, а только дополняют друг друга в процессе филогенеза.
Различие остается только в том, что функция вестибулярная есть низшая функция, филогенетически более старая, а слуховая - более новая, диференцированная, явившаяся дальнейшей эволюцией вестибулярной как в отношении развития самого лабиринта - cochleae из lagenulae, так и корковых центров слуха.
Не развивая подробно этой точки зрения, мы привели здесь основные ее положения ввиду их исключительного интереса.
(О физиологических взаимоотношениях преддверного нерва и мозжечка будет сказано ниже).
|
The requested URL /down.htm was not found on this server.