
The requested URL /topic.htm was not found on this server.
Внутри костного внутреннего уха, или костного лабиринта, находится значительно меньшее по размерам перепончатое внутреннее ухо, или перепончатый лабиринт, с шестью нервными концевыми аппаратами VIII пары головных нервов. Перепончатое внутреннее ухо представляет собою совершенно замкнутую, наполненную эндолимфой, систему каналов и полостей, непосредственно соединенных между собою, выстланных внутри эпителием, снабженных шестью концевыми нервными аппаратами и заключенных эксцентрично в костную капсулу, повторяющую его форму. В тех местах, где находятся нервные концевые аппараты, стенки перепончатого лабиринта плотно прилегают к его костным стенкам.
Остающиеся различной формы и величины пространства между перепончатым и костным лабиринтами представляют собою систему более или менее связанных между собою перилимфатических полостей с их жидким содержимым - перилимфой. Последняя находится в непосредственном сообщении через aquaeductus cochleae с субдуральным пространством, куда изливаются также лимфатические сети, окружающие нервные стволы и сосуды.
Перепончатый лабиринт, в свою очередь, также наполнен лимфоподобной жидкостью, которая по своим физико-химическим свойствам мало отличается от перилимфы. Обе жидкости (перилимфа и эндолимфа) диффундируют между собою, т.е. проникают отчасти одна в другую через перепончатую стенку, разделяющую их, и таким образом обмениваются своими составными частями. Конечно, если бы они были одинаковы по своим физико-химическим свойствам, то никакая диффузия не могла бы иметь места.
Некоторые авторы в эндолимфе находят только следы белка, а в перилимфе, напротив, больше белка (Danhardt - у рыб). Из этого заключают, что эндолимфа, вероятно, свободна от белков, а перилимфа бедна ими.
Zange, чтобы подтвердить это, указывает на полное или почти полное отсутствие осадков в этих жидкостях здорового внутреннего уха при гистологических исследованиях (при фиксации) и на то, что эти жидкости не воспринимают красок.
По Rossi, вискозность (вязкость) эндолимфы больше, чем вискозность перилимфы. По исследованиям Ascher'a, как эндолимфа, так и лерилимфа жидки и прозрачны, как вода.
Совершенно своеобразная структура и расположение нервного концевого аппарата улитки (Кортиев орган находится на свободно натянутой мембране между двумя перилимфатическими пространствами), с одной стороны, и структура и расположение концевых аппаратов преддверных мешочков и перепончатых полукружных каналов (maculae, cristae staticae на мощных утолщениях, прикрепленных к кости) - с другой, вполне соответствуют их различным, своеобразным функциям. Кроме того, топографические и гистолого-анатомические различия этих двух отделов внутреннего уха также обусловливают неодинаковую функцию их. Б то время как первый отдел (улитка) заведует слуховой функцией, второй отдел (преддверно - полукружный) обладает статико-динамической функцией или представляет собою орган чувства пространства.
Таким образом, как с чисто анатомической, так и с физиологической точек зрения, будет более целесообразным разделить перепончатое внутреннее ухо человека на преддверно - полукружный - вестибулярный отдел с его преддверием (utriculus и sacculus) и тремя полукружными каналами и на улитковый или кохлеарный отдел (Alexander).
а) Вестибулярный, или преддверно-полукружный отдел. Вестибулярный отдел перепончатого внутреннего уха с его sacculus, utriculus и тремя из него исходящими полукружными каналами образует задне-верхнюю часть перепончатого лабиринта и со своими пятью нервными концевыми аппаратами и головными центрами представляет орган чувства пространства.
Гистологическое строение ткани перепончатого вестибулярного отдела относительно просто и почти во всех своих частях одинаково. Перепончатые оболочки эндолимфатических каналов и мешочков снаружи выстланы эндотелиоидным эпителием, а костные стенки перилимфатических пространств покрыты эндостом с его эндотелиоидным эпителием.
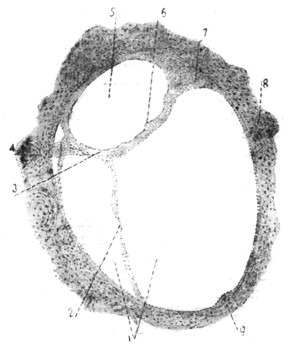 |
Рис. 14. Поперечный разрез костного и перепончатого полукружных каналов взрослого человека (увеличение в 65 раз) (по препарату Scheibe). |
В области полукружных каналов и пограничных поверхностей utriculus они соединены между собою системой соединительно - тканных, натянутых перилимфатических перемычек. После слизистого превращения и разжижения эмбрионального соединительнотканного слоя, окружающего перепончатый лабиринт, они образуют опорную сеть для него. Эта сеть состоит из соединительнотканных клеток с хорошо красящимся овоидным ядром и с многочисленными протоплазматическими отростками, которые переходят в соединительнотканные волокна, соединяющиеся с соседними клетками. Волокна, наконец, проникают, с одной стороны, в эндост, а с другой - в соединительнотканную оболочку перепончатых стенок.
В этих перемычках, также покрытых эндотелиоидным эпителием, проходят кровеносные сосуды, пересекающие, таким образом, лимфатические пространства, и здесь же встречаются иногда единичные клетки с пигментом (у человека) (рис. 14).
Сама стенка мешочков и полукружных каналов в общем очень тонка (0,5 микрон) и состоит из соединительнотканной membrana propria и однослойного эпителиального покрова. Последний в участках концевых нервных аппаратов значительно утолщается (до 1000-2000 микрон), располагаясь на сравнительно большом пространстве на бесструктурной основной мембране (membrana basilaris). Membrana propria - как субъэпителиальный слой - состоит из волокон, которые, подобно propria роговой оболочки, образуют пластинку.
Между волокнами последней встречаются звездообразные, разветвленные, анастомозирующие между собой клетки, которых в более толстых участках мембраны больше, чем в более тонких; кроме того, здесь обнаружена и крупнопетлистая сеть тонких эластических волокон. Эпителий здесь преимущественно однослойный, плоский; клетки его расположены на membrana propria - как плоские, полигональные пластинки. Исключение составляет основание utriculus, на котором клетки несколько уже и выше, а местами образуют возвышения с более высокими, цилиндрическими клетками. Так называемый raphe (шов) вдоль вогнутой поверхности полукружных каналов и их ампулл состоит внутри из эпителиальных, узких и низких клеток цилиндрической формы.
Все пять концевых нервных аппаратов окружены слоем более высокого цилиндрического эпителия, который в области трех cristae ampullares особенно ясно выступает, располагаясь вокруг их в форме planum semilunatum.
Высокий клеточный слой последнего, отделенный от самой crista узким эпителиальным желобком, к противоположному краю уплощается, постепенно переходя в простой плоский эпителий ампуллярной стенки.
Ductus endolymphaticus, отходящий от sacculus и соединяющийся узким другим каналом с utriculus, направляется к задней черепной ямке в aquaeductus vestibuli и оканчивается интрадурально в saccus endolymphaticus. Эпителий ductus и saccus endolymphaticus значительно выше, чем упомянутый индифферентный эпителий перепончатой стенки лабиринта, и имеет большое шаровидное, интенсивно красящееся ядро. Эпителий здесь расположен на базальной мембране, окруженной рыхлой соединительной тканью и волокнами твердой мозговой оболочки.
Внутри этого канала и расширения, выстланных эпителием, образуется много складок и узких длинных углублений, а к старости находят в нем целое сплетение тонких канальцев (в среднем и верхнем отделах).
Сам saccus endolymphaticus, особенно в детском возрасте, представляет собою совершенно прямую, покрытую эпителием щель в толще твердой мозговой оболочки, со складками и ворсинками на одном или на обоих концах. Величина saccus 2-4-6 мм. Эпителий его, не имея tunica propria, связан рыхло с проходящими здесь волокнами durae.
Содержимое saccus, особенно у детей, совершенно жидко и прозрачно, у взрослых же в его глубоком отделе находят аморфную, очень мелкую, суспендированную в жидкости известковую пыль.
В исключительных случаях здесь бывает слизь, но большею частью saccus представляется пустым и спавшимся.
Все пять концевых нервных аппаратов вестибулярного отдела отчасти сходны между собою по своей структуре.
Их невроэпителий, одинаковый по своему строению и совершенно изолированный от воздействия перилимфы, расположен на очень толстом нервно - соединительно - тканном слое, который, в свою очередь, прилежит к плотной основе костной лабиринтной стенки, и только в utriculus лежит на толстой соединительно - тканной пластинке, не прилегающей к кости.
Концевые аппараты, таким образом, как по своему расположению, так и по своей структуре, а также и анатомически и функционально в основном отличаются от концевого аппарата улитки (Кортиева органа).
Однако и нервные концевые аппараты преддверных мешочков, с одной стороны, и перепончатых полукружных каналов - с другой, представляют некоторые различия как в форме эпителиального слоя, так и в, высоте и в типе покровного слоя с их чувствующими волосками, передающими специфическое раздражение.
Нервный эпителий maculae (staticae) sacculi et utriculi расположен в форме овальных, несколько возвышенных пятен, величиною в 1,5- 2,0 мм, на слабо вогнутой membrana basilaris, которая своей бесструктурностью резко отличается от находящегося под нею несколько утолщенного соединительно - тканного слоя, пронизанного нервными пучками и множеством капилляров.
На этих пятнах (macula) совершенно узкий эпителий постепенно возвышается до 0,03-0,04 мм, исходя из плоского эпителия индифферентной окружности, и производит впечатление слоистого цилиндрического эпителия (при рассматривании сверху).
Однако на соответственных разрезах удается распознать, что эпителий здесь состоит из двух совершенно различных, закономерно расположенных родов клеток, а именно: 1) из нитевидных, или опорных, не имеющих никакого отношения к передаче раздражения, так как удается проследить их постепенный переход в окружающие индифферентные клетки, и 2) из волосатых или чувствующих клеток, оплетенных нервными волокнами, но не достигающих своими нижними концами до membrana basilaris. Опорные клетки проходят через всю толщу эпителия, от membrana basilaris до свободной его поверхности. В них различают нижнюю половину - как тело клетки разнообразной формы, и верхнюю, идущую' кверху между волосатыми клетками,- как нитевидный отросток.
Своими телами эти клетки выполняют все пространство, находящееся под волосатыми клетками, которым служат опорой, располагаясь на подлежащем слое, т.е. membrana basilaris, то широкими, то веретенообразными своими основаниями.
Ядра опорных клеток находятся на разных высотах, отчего на разрезах часто получается вид нескольких лежащих один над другим клеточных слоев, однако эти ядра обычно находятся под слоем волосатых клеток и всегда по размерам меньше, чем ядра волосатых клеток.
Внутри этих нитевидных опорных клеток найдены интрацеллюлярные опорные волокна (тонофибриллы), аналогичные опорным волокнам Дейтерсовых клеток Кортиева органа (Held).
Опорные волокна идут в клетке от membrana basilaris кверху и переходят на свободную поверхность кисточками в поверхностную сеть, оплетающую головки волосатых клеток. Сеть представляет собою кутикулярную мембрану, покрывающую всю поверхность maculae, аналогично macula reticularis Кортиева органа.
Волосатые, или чувствующие клетки этих концевых аппаратов приблизительно цилиндрической формы, с округленным и колбообразно расширенным нижним концом, содержат большое, богатое хроматином, ядро.
Нижние концы этих клеток не доходят до membrana basilaris, оканчиваясь на половине высоты эпителиального слоя.
 |
Рис. 15. Разрез через macula statica взрослой кошки при действии на cupula хлористым кальцием со стороны барабанной полости (для диффузии) с последующей прижизненной фиксацией (через сосуды). Примененная здесь фиксация вследствие сильных диффузионных токов устранила более тонкие волокна (волоски) и часть гомогенной массы, выполняющей cupula, так что здесь остались ясно заметными только более толстые волокна и отдельные плазматические шары. Здесь также ясно выступает сохранившаяся волокнистая сеть (сильное увеличение) (по A. Eckert - Mobius'y). |
Так как некоторые волосатые клетки не одинаковой длины, то их ядерная область может доходить до ядерной области опорных клеток. Вокруг всей поверхности волосатых клеток безмякотные нервные волокна n. vestibularis образуют густую зернистую сеть. Такая же, но более густая, сеть у основания волосатых клеток охватывает их бокалообразно снизу и подымается кверху тонкими ветвями по боковой поверхности этих клеток. Такие зернистые безмякотные нервно-волокнистые сплетения спускаются также вниз, прободают membrana basilaris и, превратившись в миэлиновые нервные волокна, наконец переходят в субъэпителиальную ткань, где образуют пучки, отходящие к более толстым разветвлениям n. vestibularis.
Верхние концы волосатых клеток оказываются заключенными в кутикулярную пластинку, расположенную над ними. В эту кутикулярную мембрану переходят и верхние концы интрацеллюлярных волокон нитевидных опорных клеток (тонофибриллы). С верхней поверхности волосатых клеток отходит кверху плотный чувствующий волосок (20-25 микрон), состоящий из многочисленных очень тонких параллельных волосков; продолжаясь кверху, они разветвляются и, переплетаясь между собою, образуют тонкую волокнистую сеть, расположенную почти параллельно верхней поверхности эпителия и своими концами переходящую непосредственно в отолитовую мембрану.
Отолитовая мембрана, по мнению многих авторов, представляет собою параллельную верхней поверхности эпителия maculae staticae пластинку, состоящую из переплетенных между собою продолжений тончайших чувствующих волосков, и образующую сеть над всею macula. Петли или ямки этой сети заполнены желеобразной массой, в которой находятся отолиты, т.е. масса кристаллических телец, белая, как мел, при падающем свете. Отолиты, или тельца - обычно продолговатой формы со слегка закругленными краями шестиугольники, но у различных животных они имеют разную форму, присущую данному животному. Состоят они, главным образом из углекислой извести и органической основы, которая и после декальцинации в большинстве случаев остается и сильно красится гемотоксилином. Отолитовый слой по своему удельному весу оказывается значительно тяжелее, чем окружающая его лабиринтная жидкость, и потому при очень быстром вращении животного может отрываться от своей основы (Wittmaack).
Пространство между отолитовой мембраной и верхней поверхностью эпителия, помимо упомянутой сети из чувствующих волосков, выполнено жидкой массой, отличающейся по своему составу от окружающей лабиринтной эндолимфы (рис. 15). Это видно из того, что при фиксации из этой промежуточной массы выпадают вещества, которые хорошо красятся, a liquor labyrintht остается бесцветной (Eckert - Mobius).
 |
Рис. 16. Crista ampullaris заднего вертикального полукружного канала новорожденного (человека). Продольный разрез. (Увеличение в 150 раз). (Микрофотограмма с препарата Н. Ф. Попова). |
Crista (statica) ampullaris, в противоположность macula statica, значительно возвышается над своим основанием и боковыми стенками ампуллы в форме узкого, выступающего в просвет ампуллы, усеченного конуса. Конус покрыт нейроэпителием, над которым находится желеобразное образование, также в виде усеченного конуса (cupula), как бы насаженное на волоски его эпителия.
Нейроэпителий cristae только немного выше эпителия maculae, но во всех своих структурных деталях, как опорных, так и чувствующих клеток с их волосками и с их иннервацией, вполне сходен с нейроэпителием maculae.
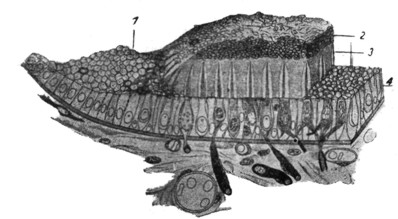 |
Рис. 17. Пластическо-схематическая реконструкция гистологических соотношений на продольно краевом разрезе maculae (по W. Kolmer'y).
|
В crista чувствующие волоски почти параллельно отходят от своих клеток вверх и только по краям несколько конвергируют и проникают в покрывающую их cupula, распределяясь в ней довольно равномерно. Отолитовая мембрана здесь отсутствует, хотя в последнее время и здесь были открыты Kubo отолиты при неполной декальцинации их.
Что касается cupula cristae ampullaris, то это образование представляется преформированным при жизни и отличающимся по своему составу от лабиринтной эндолимфы, так как при фиксации оно коагулируется в слабо красящуюся гомогенную массу (рис. 16).
Необходимо еще упомянуть о так называемой рудиментарной macula neglecta, которую Retzius нашел у целого ряда низших позвоночных, рептилий и птиц на основании utriculus, у места перехода sinus utricularis inferior в ампуллу нижнего полукружного канала.
Аналогичное образование найдено у человека (в 96% исследованных височных костей, Stutz, Benjamins, Ruttin, Sakai).
Это небольшое, неправильной формы, скорее, коническое возвышение находится в только что указанном месте или, точнее, между устьем ductus endolymphaticus и crus commune или в конце последней.
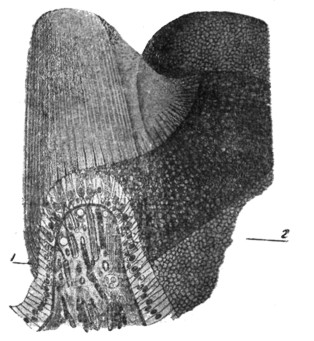 |
Рис. 18. Пластическо - схематическая реконструкция гистологических соотношений половины crista, разрезанной на две почти равные части в продольном направлении самой ампуллы (по W. Kolmer'y). |
Macula neglecta покрыта кубическим переходным эпителием или высоким цилиндрическим, но только не диференцированным на волосатые и опорные клетки, т.е. он имеет характер эпителия plani semilunati. Ни чувствующих волосков, ни cupula, ни отолитов у человека в этом образовании не находили, хотя у человеческих зародышей отмечали едва заметные рудиментарные остатки этих составных частей нормальной maculae (рис. 17 и 18).
б) Перепончатая улитка. Передне-нижняя часть внутреннего уха представляет улитку с ее концевым нервным аппаратом - Кортиевым органом.
Макроскопическая структура внутреннего уха и, в частности, улитки изложена в предыдущей главе настоящей книги. Здесь же нам предстоит коснуться только более тонкой (микроскопической) структуры улитки и упомянуть только о тех деталях, которые могут иметь непосредственное отношение как к нормальной, так и патологической физиологии этого сложного органа. Перилимфатические пространства улитки, так называемые scala vestibuli и scala tympani, с непосредственно выходящим из нее aquaeductus cochleae внутри выстланы большей частью тонким слоем эндоста, содержащего отдельные клетки с пигментом. Он распространяется также на тонкие костные пластинки laminae spiralis osseae. Эндост покрыт очень нежным эндотелием, который образует складки над более крупными эндостальными кровеносными сосудами.
Эндостальный соединительно - тканный слой выстилает все каналы и перегородки между ними также и в modiolus. Переходя на наружную, периферическую сторону завитка улитки, эндост значительно утолщается в соединительнотканный слой, снабженный многочисленными сосудами и имеющий на поперечном разрезе форму серпа - ligamentum spirale. Эта связка на всем протяжении улитки служит наружной границей обеих лестниц и самого улиткового канала.
Почти на средине (в центре) своей внутренней поверхности ligamentum spirale образует заостренный выступ, переходящий непосредственно в membrana basilaris улиткового канала. Спиральная связка состоит из почти безъядерной густой соединительной ткани, но в средних слоях она построена из рыхлой, мягкой, с многочисленными веретенообразными ядерными клетками соединительной ткани (рис. 19).
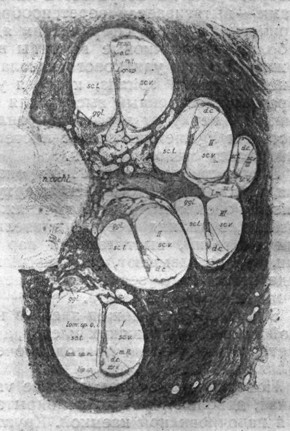 |
Рис. 19. Вертикальный разрез через ось, улитки взрослого человека при среднем увеличении (по A. Eckert - Mobius'y). |
Ductus reuniens - канал, соединяющий sacculus и ductus cochlearis, имеет очень тонкую стенку, в особенности при переходе в улитку. Структура этой стенки очень походит на структуру свободной стенки sacculus и membrana Reissneri, в которые она непосредственно продолжается. Она состоит из очень тонкого соединительнотканного слоя, покрытого внутри однослойным низким, плоским эпителием с плоскими ядрами.
Эндолимфатическое пространство улитки представляет улитковый канал, имеющий на поперечном разрезе форму треугольника. Верхнюю сторону, или вестибулярную стенку его, образует membrana Reissneri, наружную - вестибулярная половина ligamenti spiralis, или так называемая stria vascularis, и нижнюю - тимпанальная стенка, или membrana basilaris.
Верхняя вестибулярная стенка улиткового канала, или membrana Reissneri, своим внутренним нижним концом прикрепляется у начала cristae spiralis на вестибулярной поверхности laminae spiralis osseae, направляется наискось кверху и кнаружи к месту своего верхне-наружного прикрепления, к верхней части ligamenti spiralis.
Membrana Reissneri представляет собой очень нежную тонкую, бессосудистую, почти бесструктурную, соединительно - тканную пластинку, пронизанную очень тонкими эластическими волокнами.
С вестибулярной наружной поверхности она выстлана слоем эндотелия, а с поверхности, обращенной к улитковому каналу, покрыта простым полигональным плоским эпителием, в единичных клетках которого иногда встречается пигмент. Эпителий этот иногда - местами - вытягивается веретенообразно, а иногда образует круглые зернистые, с более сферическим ядром, клетки или - очень редко- небольшие сосочковые выступы в полость улиткового канала. Наружная стенка улиткового канала образуется вестибулярной половиной ligamenti spiralis, которая у начала тимпанальной половины образует заостренный выступ для прикрепления membranae basilaris. Книзу и кверху от этого прикрепления, т.е. по направлению в обе лестницы ligamentum spirale постепенно истончается, переходя в тонкий эндост стенок лестниц.
В области улиткового хода она покрыта слоем высокого эпителия, который, продолжаясь кверху, переходит в плоский эпителий Reissner'овой мембраны, а продолжаясь книзу, переходит на membrana basilaris, в цилиндрический эпителий или в клетки Claudius'a. Высокий эпителий, выстилающий наружную стенку улиткового канала или, как ее называют, stria vascularis (сосудистая полоска или лента), содержит много зернышек, и между ними встречаются зернышки темно-желтого пигмента.
Эпителий расположен здесь в несколько слоев, причем верхний слой состоит из высоких клеток, а более глубокий - из более низких клеток; между ними проходит очень много капиллярных и предкапиллярных сосудов.
Поверхность эпителия striae vascularis, обращенная в улитковый канал, довольно ровная и покрыта тонкой, кутикулярной исчерченной палочковидной каемкой. Круглые ядра этого верхнего слоя эпителия большей частью расположены ближе к свободному его краю.
Базальная же поверхность этого эпителия, имея неправильные протоплазматические отростки, образует неровную поверхность с бухтами и нишами.
В этих бухтах проходят совершенно обнаженные кровеносные сосуды (без сопровождающей соединительной ткани) и образуют здесь обильные поверхностные капиллярные сети. Таким образом, мы здесь встречаемся с эпителием, содержащим сосуды, а потому его называют сосудистым эпителием. Под ним расположен широкий полиэдрический слой эпителоидных клеток с зубчатыми отростками, которые выполняют промежутки между глубокими капиллярами, образуя сеть (ретикулярный слой) (рис. 20).
Итак, эпителий striae vascularis по своей структуре может быть признан выделительным органом, участвующим в образовании эндолимфы. Нижнюю треть striae vascularis, где выступает prominentia spiralis со своим сосудом, выстилает однослойный эпителиальный покров наружной стенки. Он состоит из более плоских, одинаково зернистых, кубических клеток, которые по направлению к sulcus spiralis externus делаются более высокими. Эпителиальные клетки sulcus spiralis externus на свободной своей поверхности и по высоте и по форме представляют собой цилиндрический эпителий, но с базальной поверхности клетки его образуют пальцеобразные разветвляющиеся отростки, которые проникают глубоко в соединительную ткань, также богатую сосудами. Таким образом, границы между этим эпителием и прилежащей соединительной тканью совершенно сглажены. Третья - тимпанальная стенка ductus cochlearis состоит из crista spiralis, расположенной на lamina spiralis, отчасти заходящей за нее, и из перепончатой мембраны, соединяющей lamina spiralis ossea с ligamentum spirale, т.е. из membrana basilaris, на внутренней части которой находится самая важная часть улитки - Кортиев орган, или papilla basilaris.
Crista (lamina) spiralis или limbus laminae spiralis представляет постепенно увеличивающееся к периферии утолщение эндоста laminae spiralis с ее вестибулярной поверхности. Это образование постепенно уменьшается и в ширине, и в толщине от основания улитки к ее вершине.
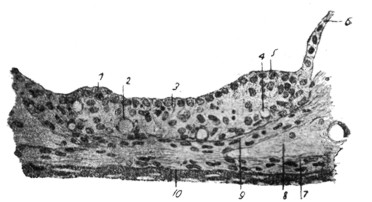 |
Рис. 20. Рздиарно - поперечный разрез через stria vascularis из верхнего завитка улитки 32-летней женщины (по Kolliker'y).
|
Limbus laminae spiralis, главным образом состоит из очень твердой, более или менее гомогенной, соединительной ткани, которая в глубоких слоях имеет радиарные волокна с ядрами и сосуды, и из эпителиального покрова в области улиткового канала.
Вестибулярная губа (lаbium vestibulare) limbus'a или клинообразный его выступ, на котором прикреплена часть membranae tectoriae, представляется на радиарных срезах совершенно гладкой. На плоскостных же срезах оказывается, что на ее стенке, обращенной к membrana Reissneri и с периферии ограниченной острым краем, верхняя поверхность только кажется гладкой, а в действительности в отделе, прилежащем к crista membranae Reissneri, на ней выделяются овальные или палочкообразные плоские выступы, разделенные бороздками (желобками). Эти радиальные бороздки, направляясь равномерно к периферии и несколько углубляясь, доходят до острого края labium vestibulare и образуют выступы, так называемые слуховые зубцы Huschke. Своими выступами, валиками или ребрами они выдаются над вестибулярной поверхностью, к которой прилежит часть membranae tectoriae, и над sulcus spiralis internus, но наиболее сильно развиты они в основном завитке улитки и значительно меньше в верхнем завитке.
Вся обращенная к membrana Reissneri свободная поверхность этой соединительно-тканной cristae spiralis с ее бороздами, валиками (ребрами) и слуховыми зубцами покрыта отчасти цилиндрическими клетками, которые в бороздах и на ребрах слуховых зубцов, делаются более плоскими и, как полигональный плоский эпителий, переходят на поверхность sulcus spiralis internus и вестибулярную поверхность labium tympanicum.
Тимпанальная губа (labium tympanicum) представляет собой два костных тонких листка: верхний - вестибулярный и нижний-тимпанальный, разделенные проходящим между ними нервным пучком.
Эти два костные листка laminae spiralis osseae со своим эндостом продолжаются значительно кнаружи, выступая много дальше, чем вестибулярная губа.
Тимпанальный листок состоит из соединительной ткани, содержащей ядра (эндоста), и покрывающего его нежного эндотелиального слоя и прилежит к месту прохождения нервов, слегка изогнувшись к вестибулярному листку. Здесь labium tympanicum и заканчивается, переходя в membrana basilaris.
Более толстый вестибулярный листок, покрытый плоским эпителием, образует базальную границу sulcus spiralis internus и в своих глубоких частях, прилегающих к нервным пучкам, снабжен веретенообразными клетками.
Его периферическая часть (habenula perforata) имеет целый ряд расположенных спирально радиальных продолговато - овальных отверстий (foramina nervosa), через которые входят в Кортиев орган, в косом направлении, волокна п. cochlearis. Из соединения обоих листков тимпанальной губы (labium tympanicum) исходит membrana basilaris и, сильно истончаясь, направляется прямо к заостренному выступу ligamenti spiralis. В membrana basilaris различают: 1) внутренний отдел - zona arcuata, покрытый частью Кортиева органа (дугами) и простирающийся до наружного конца ножки наружной Кортиевой дуги; 2) средний отдел - zona tecta, занятый остальной частью Кортиева органа и продолжающийся кнаружи до последней клетки Hensen'a, и 3) наружный отдел - zona pectinata membranae basilaris, переходящий в ligamentum spirale. Основная перепонка состоит из трех слоев: 1) собственного - волокнистого, 2) вестибулярного - пограничного и 3) тимпанального -обкладочного слоя.
Волокнистый слой состоит из очень тонких, густо расположенных, крепко спаянных между собою, радиально направленных фибрилл, которые представляют продолжение тонких волокон - labium tympanicum.
В zona arcuata (отчасти), в zona tecta и в zona pectinata они превращаются в более толстые, слегка уплощенные с боков, не ветвящиеся волокна или в акустические (слуховые) струны, которые на уплотненных препаратах можно легко измерить, но только спаянные между собою.
Подходя к ligamentum spirale, струны переходят в ее волокна, которые отчасти направляются радиарно, но преимущественно загибаются по направлению как к scala tympani, так и к scala vestibuli. Очень тонкий и почти гомогенный вестибулярный пограничный слой, в который переходит без резкой границы интерфибриллярная склеивающая субстанция под ним лежащего волокнистого слоя, содержит местами радиально расположенные клеточные ядра; в zona tecta их нет.
По направлению к ligamentum spirale вестибулярный слой кажется изрешетенным, что можно проследить на некотором расстоянии под эпителием наружной стенки улитки. В наружном отделе membranae basilaris он покрыт цилиндрическим эпителием, или клетками Claudius'a.
Более толстый тимпанальный, обкладочный слой представляет собою, главным образом, продолжение тимпанального листка labium tympanicum. Co срединным волокнистым, над ним лежащим слоем: он соединен гомогенным склеивающим веществом.
Этот тимпанальный обкладочный слой также можно рассматривать как остаток соединительной зародышевой ткани scalae tympani. В средине сама membrane basilaris представляется наиболее толстой, а в наружной части к ligamentum spirale она постепенно делается более тонкой.
Кортиев орган, или papilla spiralis, papilla acustica basilaris, представляет собою концевой нервный аппарат улиткового нерва. Как показывает история развития, Кортиев орган происходит от своеобразного превращения эпителиальных клеток и их кутикулярных образований, расположенных на membrana basilaris в форме спирально идущего через все завитки улитки эпителиального, сравнительно широкого, валика различной высоты.
Как в начале основания улитки, так и в вершине ее Кортиев орган, как таковой, совершенно отсутствует, вместе с тем в этих концевых частях отсутствует и membrana basilaris.
Опорные клетки Кортиева органа и их кутикулярные покровные пластинки представляют очень сложно устроенный опорный аппарат для волосатых клеток Кортиева органа. Этот аппарат, прежде всего, состоит из двух групп специфических опорных клеток с образованными внутри их интрацеллюлярными опорными волокнами (тонофи-бриллами) и из поверхностных, кутикулярных, обхватывающих колец для закрепления головок волосатых клеток. К специфическим опорным клеткам относятся Кортиевы дуги и Дейтерсовы клетки, к которым Held причисляет так называемые внутренние фаланговые клетки и внутренние пограничные клетки.
К неспецифическим опорным клеткам относятся так называемые внутренние опорные клетки и клетки Hensen'a.
Эти последние клетки, подобно высокому переходному эпителию, вокруг macula и crista statica можно рассматривать как общий слой клеток, откуда происходят специфические диференцированные, т.е. опорные и волосатые клетки, поэтому их правильнее назвать, как предлагает Eckert-Mobius, клетками переходного типа.
Самые прочные опорные образования, значительно отличающиеся от обыкновенной формы эпителиальных клеток, представляют собою так называемые Кортиевы дуги, которые образуют два спирально идущих ряда: внутренний и наружный.
Дуги (клетки), соединяясь между собою головками и расходясь основаниями, образуют (на поперечном разрезе) треугольник, а вся система дуг представляет интраэпителиальное пространство, или туннельное пространство, проходящее спирально через все завитки улитки.
Внутренние Кортиевы дуги своими расширенными основаниями или протоплазматическими ножными пластинками, содержащими ядро, расположены на membrana basilaris к периферии от foramina nervosa. Эти ножные пластинки распространяются по основанию туннеля кнаружи и своими заостренными концами вклиниваются между тонкими же чередующимися ножными пластинками наружных Кор-тиевых дуг. Таким образом, внутренние и наружные Кортиевы дуги своими ножками занимают все основание туннеля. Ножная пластинка каждой внутренней Кортиевой дуги переходит кверху в продольную стройную среднюю часть, которая заканчивается верхней, несколько утолщенной головкой. С внутренней стороны последней выступает
короткий внутренний отросток, внедряющийся между внутренними волосатыми клетками. На наружной стороне головки имеется сочленовное углубление для головки соотнетствующей наружной Кортиевой дуги, а над этим углублением и этой головкой находится покровная (кутикулярная) продолговатая прямоугольная головная пластинка. Своими кутикулярными, слегка вогнутыми краями она отграничивает внутренние волосатые клетки с одной стороны, а своим почти прямолинейным, наружным краем доходит до первого ряда наружных волосатых клеток. Внутри Кортиевых дуг, в особенности при продолжительной фиксации хромовыми солями, выявляется система окрашенных опорных волокон, или тонофибрилл, в щелочах легко растворимых.
Во внутренней части ножной пластинки внутренней дуги эти опорные волокна в большом количестве доходят до membrana basilaris и прикрепляются к ней, образуя конус вершиной кверху и конусообразное тельце (опорный конус). В средней части дуги эти тонофибриллы сближаются в более тонкое стержнеподобное волокнистое образование; волокна его, подойдя к верхней части дуги, опять расходятся в головное утолщение. Своими верхними концами они прикрепляются отчасти к вогнутой наружной поверхности головки, отчасти теряются во внутреннем отростке. Однако большая часть этих концов расходится дугообразно, направляясь кнаружи в головную пластинку, и оканчивается на ее периферическом краю, прилежащем к первому ряду наружных волосатых клеток (Held, Kolmer).
Наружных Кортиевых дуг меньше, чем внутренних, и потому две-три наружные Коотиевы дуги приходятся на одну внутреннюю. Наружные дуги имеют S-образно изогнутое тело и такую же ножную пластинку, как и внутренние дуги. Эти пластинки, содержащие ядро, своей протоплазматической ножкой направлены обратно, т.е. кнутри от пучка опорных волокон почти до соприкосновения с заостренным краем пластинки внутренней дуги.
Головной конец наружной дуги своей выпуклостью - сочленовной головкой, направленной кнутри, входит в углубление головки внутренней дуги, причем они спаиваются неподвижно, так что здесь не может быть речи о суставном соединении. От наружного конца головки наружной дуги под головной пластинкой внутренней дуги отходит фаланговый отросток и оканчивается веслообразно расширенной пластинкой - фланговой пластинкой первого ряда. Вогнутые длинные ребра этих пластинок, прилегающие одна к другой, образуют между собой круглые отверстия, окаймленные кутикулярными, обхватывающими кольцами (кольцевыми фасетками) для верхвих концов (головок) наружных волосатых клеток I ряда.
Через среднюю часть наружной Кортиевой дуги также проходит пучок опорных волокон (тонофибрилл) (около 27 - Kolmer), которые в наружной части нижней пластинки расходятся для обхватывания конусообразного базального тельца и прикрепляются к membrana basilaris. Эти волокна, дойдя до верхнего конца Кортиевой дуги, опять расходятся, образуя коническое расширение, оканчивающееся в головке внутренней дуги, и тут на ее поверхности образуют кутикулярное утолщение в форме пластинки.
Кнаружи от Кортиевых дуг в Кортиевом органе расположены 3 или 4 чередующихся ряда другого типа специфических опорных клеток, так называемых Дейтерсовых клеток. Цилиндрические тела
этих клеток, несколько наклоненные кнутри, своими нижними концами прикрепляются к membrana basilaris, причем шестиугольные ножки первого ряда соприкасаются непосредственно с протоплазматическими ножками наружных Кортиевых дуг.
Дейтерсовы клетки имеют округлое большое ядро, расположенное на значительной высоте, и слегка зернистую протоплазму, в которой вверху клетки находится тельце Retzius'a.
Верхняя часть их поверхности, обращенной к Кортиевым дугам, превращена в головкообразный выступ тела клетки или в так называемую нижнюю головку Дейтерсовой клетки (Held). Нижняя головка Дейтерсовой клетки представляет собою базальную опорную поверхность для соответственной волосатой клетки и, кроме того, образует по оси этой поверхности щелеобразное углубление, которое, как опорный бокал, вполне обхватывает основание наружной волосатой клетки. Приблизительно на средине высоты Кортиева органа каждая Дейтерсова клетка суживается в форме косо расположенной шейки, переходящей кверху в тонкий фаланговый отросток. Отросток, изгибаясь, поднимается спирально и заканчивается бисквитообразной головной пластинкой, закрепленной между соответствующими рядами верхних концов волосатых клеток. Самый наружный ряд этих головных пластинок представляет наружный край кутикулярной рамки, образовавшейся на верхней поверхности Кортиева органа.
Через тело Дейтерсовых клеток по всей их длине проходит такой же пучок опорных волокон, как и в Кортиевых дугах, и особенно ясно он выражен в основном завитке. Эти опорные волокна отходят из центра основания клетки узким фибриллярным концом и волокнистым пучком направляются к средине клетки, где приблизительно на высоте ядра пучок расщепляется на две части. Одна из этих частей, изгибаясь, направляется спирально в вышеупомянутый фаланговый отросток и своим расширением образует здесь кутикулярную фасетку для фаланговой пластинки. Другая часть этих волокон, или другой волокнистый пучок, распадается и, подойдя к нижней головке Дейтерсовой клетки, проникает в нее и участвует в построении бокаловидного образования (Held).
Между фаланговыми отростками Дейтерсовых клеток и волосатыми клетками остаются свободные промежутки, выполненные эндолимфой. Они называются Nuel'евыми пространствами. Вследствие наклонения Дейтерсовых клеток к наружным Кортиевым дугам, между этими дугами и первым рядом Дейтерсовых клеток получается еще большее пространство, которое соединяется с туннельным через щелевые промежутки между наружными Кортиевыми дугами. За Дейтерсовыми клетками кнаружи следует 5-8 рядов клеток Hensen'a, очень богатых водой, а потому трудно фиксируемых. Эти клетки образуют широкий вал, более плоский в основном завитке улитки и более выдающийся в верхнем завитке, вследствие чего здесь образуется кнутри вдающийся угол с примыкающими клетками Claudius'a. Клетки Hensen'a своими суженными ножками прикрепляются к membrana basilaris и, расширяясь кверху, принимают несколько наклонное положение к основной мембране, а на радиальном срезе кажутся наслоенными одна на другую.
Их круглые ядра расположены на различных высотах внутри расширенных верхних половин клеток.
Светлая, мелкозернистая протоплазма их содержит - в норме - маленькие жировые зернышки, которые у грызунов (морских свинок) имеются в большом количестве; эти зернышки очень велики и при: экспериментально - гистологических работах нередко принимаются за патологическое образование.
На своей свободной верхней поверхности клетки Hensen'a покрыты кутикулярной, сравнительно толстой мембраной; в целом ее можно рассматривать как продолжение поверхностной рамки Кортиева органа.
За клетками Hensen'a следует однослойный покровный цилиндрический и отчасти кубический эпителий, или клетки Claudius'a, по структуре своей протоплазмы очень сходные с клетками Hensen'a. Они выстилают наружную часть zona pectinata и переходят на начало sulcus spiralis externus.
Этим мы заканчиваем изложение очень сложного с механической и физической точек зрения органа, являющегося опорным аппаратом для необыкновенно нежных, высокодиференцированных, и потому очень лябильных, волосатых клеток, этих специфических слуховых элементов.
Остается сказать о том, как укреплены внутренние волосатые клетки.
По последним исследованиям Held'a и Kolmer'a головки внутренних волосатых клеток также окружены кутикулярными ободками, которые исходят, главным образом, от головной пластинки и внутренних отростков внутренних Кортиевых дуг и ими поддерживаются. Рядом с верхним отделом боковых наружных половин внутренних волосатых клеток расположены со своими узкими ножками стройные тела внутренних фаланговых клеток Held'a треугольной формы, головная часть которых переходит в узкую внутреннюю фаланговую пластинку. Здесь еще имеются внутренние пограничные клетки, плотно прилегающие к внутренним поверхностям внутренних волосатых клеток.
Их ядра находятся ниже средины клетки и не имеют никаких признаков фибриллярного опорного скелета, но в средней части у них наблюдается небольшое утолщение; значительно выдаваясь, оно может служить нижней опорой для внутренних волосатых клеток. При удачной фиксации верхние отделы этих клеточных групп, окружающих внутренние волосатые клетки у человека, ясно распознаются, а средние и нижние части их почти не заметны. Вероятно, зернистость нижних отделов этих клеток растворяется, а в верхних отделах, окружающих основания волосатых клеток, она сохраняется, почему они и считаются опорными клетками для внутренних волосатых клеток (Eckert-Mobius).
По направлению к оси улитки, т.е. кнутри, к этим опорным клеткам прилегают широкие кубические эпителиальные клетки, за которыми следуют более плоские клетки, выстилающие sulcus spiralis internus.
Волосатые клетки Кортиева органа - эти специфические слуховые элементы - необыкновенно нежны, богаты водой и очень лабильны вследствие особенной физико-химической структуры их протоплазмы. Поэтому, при действии фиксирующих веществ они легко сморщиваются, но головные укрепленные концы их оказываются более устойчивыми. Так что эти клетки на гистологическом препарате представляются большею частью суженными и укороченными, нередко даже отделившимися от своих базальных опорных бокалов (что иногда принимается за прижизненные патологические изменения).
Волосатые клетки расположены параллельными спиральными рядами и делятся на внутренние и наружные волосатые клетки. Наружные волосатые клетки располагаются с наружной стороны наружных Кортиевых дуг в три ряда в основном завитке, в четыре ряда в среднем и верхнем и даже в пять рядов в конце верхушечного завитка. Как более устойчивые, сравнительно с внутренними, они представляются довольно правильными, сравнительно высокими цилиндрическими клетками, которые своим закругленным нижним концом помещаются в упомянутых опорных бокалах, плотно прилегая к их стенкам, т.е. расположены на нижних головках, находящихся на внутренней стороне Дейтерсовых клеток. Эти нижние концы волосатых клеток не связаны органически с дном бокала (рис. 21).
Из своих бокалов волосатые клетки, слегка наклонившись к оси улитки, свободно проходят через пространства Nuel'я к расположенным над ними круглым отверстиям laminae reticularis, образовавшимся из спаявшихся фаланговых пластинок.
Головки волосатых клеток, проникнув в эти кутикулярные отверстия (кольца), плотно прикрепляются к их внутренним краям.
На хорошо хромированных и окрашенных гематоксилином препаратах в диффузно - зернистом теле наружных волосатых клеток ясно видна фибриллярная структура, с которой связано находящееся в основании клетки темноокрашенное тонковолокнистое зернистое образование, или тельце Retzius'a.
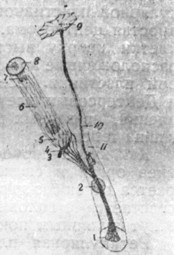 |
Рис. 21. Дейтерсова опорная клетка с наружной волосатой клеткой. Полусхематически (по Schaefer'y). |
Над этим тельцем, в нижней половине клетки, расположено большое круглое ядро, а в головном конце клетки еще находится окрашенное тельце Hensen'a, которое у человека представляется то коротким конусом, направленным вершиной вниз, то большим круглым телом с несколькими разветвленными книзу отростками.
По новейшим исследованиям (Held) это тельце Hensen'a является волокнистым включением, как продукт фиксации, состоящим из отдельных волоконец, отчасти связанных между собою, отчасти идущих параллельно или спирально. Сама головка наружной волосатой клетки, заключенная в кольце laminae reticularis, сверху покрыта нежной кутикулярной покровной пластинкой; у наружного края этой пластинки заметна простая центрозома, а в средней зоне возвышается короткий пучок заостренных волосков.
Внутренние волосатые клетки, располагаясь с внутренней стороны внутренних Кортиевых дуг одним спиральным рядом во всех завитках улитки, и по форме и по своей структуре менее вообще устойчивы, чем наружные.
У человека они очень редко встречаются хорошо фиксированными. Нередко распавшиеся пограничные базальные клетки, приставшие к нижним концам, ошибочно принимали за дендровидные протоплазматические отростки этих внутренних волосатых клеток.
Их несколько наклонное кнаружи сравнительно толстое тело в своем нижнем, более округлом, конце содержит большое ядро. Снаружи внутренние волосатые клетки ограничены внутренними фаланговыми клетками, а изнутри - внутренними пограничными клетками, которые своими выступами по средине, на половине высоты Кортиевых дуг, образуют опорную часть для них. Клеточных включений, аналогичных тельцам Retzius'a и Hensen'a, во внутренних волосатых клетках не удалось обнаружить. Их верхняя, несколько суженная и как бы косо обрезанная головная часть укреплена, главным образом, в овальном краевом промежутке, прилегающем к внутренней Кортиевой дуге. Кроме того, головная часть покрыта тонкой кутикулярной пластинкой; на наружном крае этой пластинки отмечается простая центрозома. На головной пластинке внутренней волосатой клетки кверху выступают, в два-три ряда чувствующие волоски, расположенные по всей длине длинного поперечника головной покровной пластинки. Все верхние головные части как волосатых, так и Дейтерсовых клеток и Кортиевых дуг имеют целую систему укреплений, связанных в одну решеткообразную пластинку, называемую lamina reticularis.
Как кутикулярное образование lamina reticularis, покрывающая Кортиев орган и связанная с элементами, входящими в него, простирается от sulcus spiralis internus до sulcus spiralis externus. Кнутри она срастается с головными пластинками Кортиевых дуг, а кнаружи - с кутикулярным покровом клеток Hensen'a.
Ретикулярная пластинка состоит из прозрачных нежных дву-контурных сравнительно толстых волокон, образующих петли (отверстия - кольца) разной величины. Эти отверстия - кольца служат для укрепления головок волосатых клеток. Промежутки между этими отверстиями выполнены кутикулярными бисквитообразными пластинками, похожими на фаланги пальцев и называемыми поэтому фалангами. Последние представляют собою расширения верхних концов фаланговых отростков Дейтерорвых клеток (рис. 22).
Эпителий sulcus spiralis internus непосредственно переходит в поддерживающие внутренние клетки, а к ним волнистой линией прилегает один ряд отверстий - колец для головок внутренних волосатых клеток. Наружные контуры этих клеток соприкасаются с головными пластинками внутренних Кортиевых дуг.
Кольца - отверстия для наружных волосатых клеток и фаланги чередуются между собою - кнаружи. В самых же наружных отделах форма фаланг делается неправильной, и они постепенно переходят в неправильные кутикулярные многоугольники, покрывающие клетки Hensen'a.
Над Кортиевым органом нависает - в форме продолговатой подушки - образование, которое авторы считают органом гомологичным и морфологически и эмбриологически cupula преддверно-полукружного отдела. Это-membrana tectoria, или membrana Corti. Она представляет собою перепончатое, мягкое, эластическое, обильно пропитанное жидкостью волокнистое образование. На ней можно различить более толстую (на продольном разрезе) веретенообразной формы наружную часть и - как ее продолжение - более тонкую, направленную к modiolus, внутреннюю часть, прикрепленную на вестибулярной поверхности cristae spiralis и оканчивающуюся заостренным краем у места прикрепления membranae Reissneri.
Наружная часть, соответствующая самой покровной пластинке, продолжается внутри эндолимфатического пространства только до средины Кортиева органа.
Приблизительно на средине ее плоской тимпанальной поверхности, обращенной к Кортиеву органу, выделяется выступ, переходящий потом в желобок, обозначаемый полоской Hensen'a.
Заостренный периферический край Кортиевой мембраны оканчивается коротким сетеобразно-разветвленным краевым сплетением, которое не всегда правильно расположено и очень часто завертывается кверху.
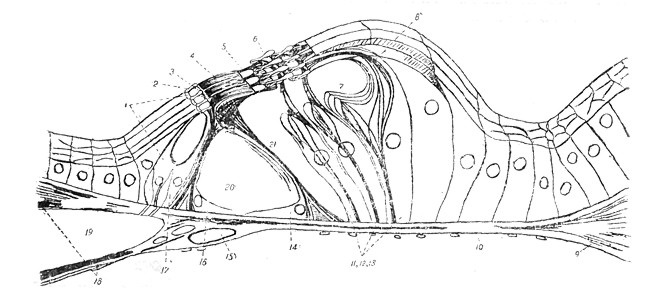 |
Рис.22. Схема строения Кортиева органа. Полу схематическое изображение всего опорного аппарата и волосатых клеток Кортиева органа (но Held'у с дополнениями Н. Ф. Попова). |
При обычных методах фиксации наружная часть Кортиевой мембраны иногда может прилегать к волоскам чувствующих клеток Кортиева органа, но на большинстве препаратов она только нависает на различной высоте - без всякого соединения с нижележащими частями. У некоторых животных она часто бывает сильно отклонена в вестибулярную сторону под углом с внутренней ее частью (некоторые экспериментаторы такое положение merabranae tectoriae принимали за прижизненное патологическое изменение).
Однако у взрослых нередко удавалось наблюдать слизистоподобные волокна и тяжи между тимпанальной поверхностью мембраны и опорными и реже - чувствующими клетками Кортиева органа.
В отношении более тонкой структуры в membrana tectoria, по Held'y, различают три слоя: средний - фибриллярный (формирующий) слой и два пограничных (поверхностных) - тимпанальный и вестибулярный, или верхнюю покровную сеть и тимпанальный пограничный слой. Фибриллярный средний слой состоит из тонких, дугообразно проходящих фибрилл и очень бледно красящейся бесструктурной межуточной субстанции. Эта последняя в наружной части (зоне) membranae tectoriae находится в большем количестве, чем во внутренней, почему во внутренней волокна отстоят одно от другого дальше и выступают яснее.
Верхняя пограничная, покровная сетка membranae tectoriae чрезмерно натянута на выпуклую вестибулярную поверхность фибриллярного среднего ее слоя. Она состоит из неодинаково сформированной сети волокнистых пучков, которые представляются сравнительно толстыми в верхушечном завитке, в краевом участке наружной части мембраны, но в других частях они значительно тоньше. Крайне нежный тимпанальный пограничный слой состоит из тонкой гомогенной нежной субстанции. На нем в области внутренней части (зоны) заметны зубчатые эпителиальные отпечатки верхней поверхности labiura vestibulare.
Здесь этот нижний слой, вероятно, приклеивает фибриллярный слой к подлежащему эпителиальному слою. В наружной части тимпанального слоя, в области петлеобразных пучков, имеются отверстия, на которые Wittmaack смотрит как на входные ворота системы соковых канальцев, имеющих связь с сосудами и с жидкостью самой membranae Corti.
По данным исследования оказывается, что в эмбриологической жизни между Кортиевым органом и наружной третью membranae tectoriae существует органическая связь, которая к концу развития делается все более и более рыхлой. Некоторые авторы указывают, что эта связь после рождения исчезает, т.е. происходит совершенное отделение membranae tectoriae от Кортиева органа. Другие авторы того мнения, что это разъединение происходит после смерти вследствие сморщивания структурных элементов.
На основании более новых исследований склонны принимать, что вся membrana tectoria в развитом ухе представляется отделенной от Кортиева органа, хотя относительно встречающихся отдельных нитевидных соединений с Кортиевым органом еще не пришли к окончательному заключению (рис. 23).
Что касается иннервации Кортиева органа, то она очень сложна и своеобразна.
Из толстого пучка нервных волокон (сплетения) n. cochlearls, находящегося между двумя пластинками laminae spiralis osseae, к наружному краю labium tympanicum отходят более тонкие пучки нервных волокон и, дойдя до habenula perforata ее, теряют свою миэлиновую оболочку, потом конусообразно суженными концами проходят по направлению к Кортиеву органу. Здесь, внутри многочисленных интраэпителиальных пространств, нервные осевые, большею частью варикозные и зернистые, цилиндры разветвляются в спиральном и радиальном направлениях, образуя несколько параллельных спиральных нервных пучков (сплетений), и, наконец, как тончайшие концевые веточки подходят к чувствующим (волосатым) внутренним и наружным клеткам и вокруг их основания образуют бокалообразные нервные сплетения.
На радиальных разрезах удается наблюдать, как у нервных отверстии habenulae perforatae входящие пучки, разветвляясь, направляются к оси улитки от внутренних дуг и подходят к внутренним волосатым клеткам через интраэпителиальные щели, между узкими ножками фаланговых и пограничных клеток, образуя здесь рыхлое нервное сплетение (внутреннее нервное, сплетение).
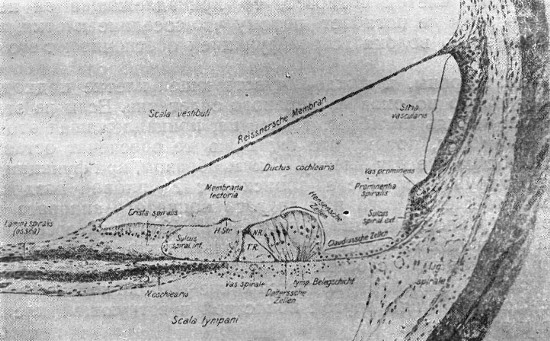 |
Рис. 23. Радиарный разрез через ductus cochlearis взрослого человека. (Оригинальный препарат из Иенской коллекции). |
Часть волокон этого сплетения идет в спиральном направлении, другая часть направляется между телами опорных клеток и основаниями внутренних волосатых клеток. Главная же масса его волокон проходит между внутренними Кортиевыми дугами и образует здесь, на их наружной стороне, над ножными пластинками в туннельном пространстве второе спиральное сплетение, или нервный туннельный компактный пучок, который тянется вдоль всего туннеля. От этого туннельного пучка отходят в поперечном направлении довольно правильными рядами через туннель кнаружи то тонкие, то толстые варикозные нервные пучки. Эти пучки между наружными Кортиевыми дугами доходят до Nuel'евых пространств и здесь - на внутренней поверхности первого ряда Дейтерсовых клеток загибаются в спирально идущий нервный пучок, или в первое наружное спиральное сплетение. С этим первым наружным нервным пучком через короткие радиарные пучки, проходящие в наружных частях Nuel'евых пространств, связаны II, III, IV и V наружные сплетения, которые располагаются возле следующих рядов Дейтерсовых клеток, аналогично первому спиральному пучку, уменьшаясь в толщине кнаружи. От каждого из этих наружных нервных спиральных пучков отходят по два или по три терминальных нервных волокна, которые подходят к основанию волосатых клеток. К внутренним волосатым клеткам они проникают между внутренними фаланговыми и пограничными клетками. Оснований наружных волосатых клеток они достигают через щелеобразную наружную вырезку в стенке опорного бокала нижней головки Дейтерсовых клеток.
Дойдя до основания волосатых клеток, нервные волокна образуют густое разветвляющееся бокалообразное Кортиево сплетение (нервный бокал), и его тончайшие, богатозернистые варикозные волоконца плотно прилегают к нижнему полюсу волосатых клеток своими конично утолщенными и богатыми нейрозомами осево-цилиндрическими ножками (концевыми сплетениями), но не проникают в протоплазму клетки (Held).
По исследованиям же Kolmer'a и Bruhl'я, нервные волоконца из этого нервного бокала проникают внутрь самой волосатой клетки и в ее верхней половине образуют тонкое сплетение из нейрофибрилл, доходящее до головного конца ее, но не имеющее связи ни с тельцем Hensen'a, ни с волосками клетки.
Таково же окончание нервов и в чувствующих клетках maculae и cristae staticae.
Остается указать на принцип (закон) иннервации концевых аппаратов перепончатого внутреннего уха. Что касается преддверно-полукружного перепончатого аппарата, т.е. macula utriculi, macula sacculi и трех cristae ampullares, то гистологические исследования (Kolmer) доказали, что концевое нервное волокно, подходя к чувствующей клетке этих нервных концевых аппаратов, разветвляется и своими нервными веточками снабжает не одну, а несколько рядом расположенных соседних чувствующих клеток. Таким образом, если какая-нибудь из этих клеток погибает, то принадлежащее ей нервное волокно при этом не погибает, потому что соседние клетки, иннервируемые от этого волокна, своей функцией обеспечивают его существование.
С другой стороны, к каждой чувствующей клетке подходит не одно, а несколько волокон из нервного сплетения. Если из этих волокон какое-нибудь погибает, то клетка, принадлежащая этому волокну, не погибает, потому что функцию ее обеспечат оставшиеся волокна, снабжающие эту клетку. Таким образом, при функции преддверно-полукружного отдела происходит групповое раздражение от нескольких клеток того или другого концевого аппарата.
Совершенно иной принцип в иннервации Кортиева органа. Здесь к каждой волосатой клетке подходит только одно концевое нервное волокно и к соседним волосатым клеткам не дает ответвлений.
В этом случае, если перерождается какое-нибудь нервное волокно, принадлежащее той или другой волосатой чувствующей клетке, то погибает и эта клетка.
И наоборот, если погибает какая-нибудь волосатая клетка Кортиева органа, то погибает и принадлежащее ей нервное волокно.
Пучок нервных волокон, выйдя из lamina spiralis и направляясь к modiolus, к главному стволу n. cochlearis, прерывается на этом пути ганглиозным узлом, или ganglion spirale.
Nervus vestibularis прерывается в meatus acusticus internus таким же узлом ganglion vestibulare, labyrinthicum или ganglion Scarpae.
Структура двух ганглиозных клеток этих двух узлов почти одинакова.
В противоположность униполярным ганглиозным клеткам спинальных ганглиев кохлеарный и вестибулярный ганглии состоят из биполярных ганглиозных клеток, из нервных волокон, исходящих из них в периферическом и в центральном направлениях, и из двух соединительно-тканных оболочек, содержащих ядра. Сами ганглиозные клетки большею частью веретенообразной или овальной формы с большим светлым пузырькообразным ядром и с равномерно распределенной зернистостью Nissl'я в протоплазме. Эти клетки переходят как в центральные, так и в периферические отростки, исходящие из противоположных полюсов их.
По Wittmaack'y миэлиновая оболочка и кератиновая сеть нервного волокна распространяются и на ганглиозную клетку, чего не бывает в спинальных ганглиозных клетках.
Bruhl'ю удалось в ганглиозных клетках gangl. spirale и gangl. vestibulare выявить целую фибриллярную систему, идущую без перерыва от одного полярного отростка до другого.
Структура нервных волокон n. octavi обычная, т.е. они состоят из нервных волоконец с ясной гомогенной миэлиновой оболочкой, прерываемой перетяжками или перехватами Ранвье, и с содержащей ядра Шванновой оболочкой.
В корешках n. cochlearis, большею частью при входе его во внутренний слуховой проход, как правило, приходится наблюдать под микроскопом то большие, то меньшие светлые зоны, в которых, особенно в их краевых участках, у более старых индивидуумов усматриваются округлые, слоистые образования, похожие на corpora amylacea.
По исследованиям Alexander'a и Obersteiner'a, эти светлые зоны п. cochlearis трактуются, как нормальные явления, подобные тем, какие наблюдаются в задних корешках спинного мозга, в которых образуется glia septum (глиозная перегородка).
Такие же глиозные пластинки, но более густо расположенные, проникают и в п. cochlearis далеко к периферии и отграничивают центральную его часть от периферической.
В этом septum, как во всех глиозных образованиях, с летами могут появиться corpora amylacea.
Такая структурная особенность присуща только чувствующим корешкам с типом центрального строения, но не моторным, где наблюдается структура, подобная периферическим нервам.
в) Нервы внутреннего уха. Восьмая пара черепных нервов иннер-вирует внутреннее ухо.
Периферические окончания этого нерва представляют собою очень сложно и своеобразно построенные концевые аппараты соответственно различным функциям уха.
N. octavus выходит из продолговатого мозга, почти рядом с n. facialis, позади оливы у заднего края Варолиева моста, двумя корешками: передним - radix vestibularis и задним - radix cochlearis.
Ствол n. VIII на своей верхне-передней поверхности образует борозду-жолоб для п. facialis, с которым соединяется посредством тончайших волоконец - fila anastomotica.
Окруженный, покрытый твердой и паутинной оболочками, как влагалищами, п. octavus вместе с n. facialis и с n. intermedius, сопровождаемый при этом arteria auditiva и vena auditiva, входит в meatus acusticus internus. Здесь же, еще до разделения, отходит от него веточка - n. ampullaris posterior и через foramen singulare направляется к ампулле заднего вертикального полукружного канала.
Отдав эту веточку, n. octavus (корешковый отдел его) делится на n. (radix) vestibularis (labyrinthicus) и n. (radix) cochlearis.
Radix vestibularis еще в meatus acusticus internus переходит в ganglion vestibulare (labyrinthicum) Scarpae по аналогии со спинальными корешками.
Этот ганглий по своей величине и по своему желтоватому цвету распознается и невооруженным глазом; на нем заметна также борозда, разделяющая его, но не вполне, на две части.
N. vestibularis, пройдя через этот ганглиозный узел, делится на две ветви: 1) ramus utriculo-ampullaris (superior) и 2) ramus saccularis (medius).
Ramus utriculo-ampullaris (superior) через мельчайшие отверстия в area vestibularis дна или основания внутреннего слухового прохода вступает в преддверие и, отдав тоненькую нервную веточку sacculus, направляется одной частью волокон к crista ampullaris горизонтального (наружного) полукружного канала - ramus ampullaris externus и к crista ampullaris верхнего полукружного канала - ramus ampullaris superior и снабжает своими нервными веточками концевые аппараты этих органов.
Другая ветвь n. vestibularis - ramus saccularis (medius) через отверстия в area vestibularis inferior дна слухового прохода направляется также в преддверие и снабжает своими разветвлениями macula sacculi, т.е. концевой аппарат преддверного круглого мешочка.
Все эти вестибулярные ветви, подходя к своим концевым аппаратам, разветвляются веерообразно и при прохождении через базальную (основную) перепонку теряют свою миэлиновую оболочку и потом проникают между опорными и чувствующими клетками, своими концевыми волоконцами образуя вокруг них бокаловидную сетку, или корзинку. Пучки волокон более мощного нервного корешка n. (radix) cochlearls, не разветвляясь, входят в tractus spiralis foraminosus и через примыкающие к нему canales longltudinales modioli подходят к широкому canalis spiralis modioli Rosenthali, или canalis ganglionaris, у места прикрепления к нему laminae spiralis; здесь, при входе в него, отделившиеся от него нервные пучки прерываются ганглиозными клетками - ganglion spirale, который представляет собою нервную округлую пластинку, идущую спирально от основного завитка до вершины улитки.
Выходящие из ганглиозных клеток нервные волокна направляются в lamina spiralis ossea, располагаются здесь между ее двумя костными пластинками и образуют сплетение пучков нервных миэлиновых волокон, которые отсюда направляются частью спирально, частью радиально.
Пучки, идущие в спиральном направлении и перекрещивающиеся с радиальными волокнами, распространяются по всему соответственному завитку (Held).
Радиальные пучки, лишившись своих миэлиновых и Шванновых оболочек, проходя через нервные отверстия habenula perforate, направляются прямо к основанию чувствующих (волосатых) клеток Кортиева органа и здесь оканчиваются, как выше было описано (рис. 24).
г) Кровеносные сосуды перепончатого внутреннего уха происходят из arteria auditiva (ветвь arteriae basilaris, образующаяся из двух arteriae vertebrales), которая вместе с п. VIII и п. VII входит во внутренний слуховой проход и здесь делится на: 1) arteria vestibuli и 2) arteria cochleae communis.
Arteria vestibuli проходит вместе с n. utriculo-ampullaris и снабжает густой капиллярной сетью macula utriculi и crista ampullaris верхнего и наружного полукружных каналов. Далее своими петлистыми разветвлениями она распространяется на начала полукружных каналов и на верхние части utrlculus и sacculus.
Arteria cochleae communis делится на: 1) arteria vestibule-cochlearis и 2) arteria cochlearis propria.
Arteria vestibule - cochlearis отдает одну ветвь, которая направляется к crista ampullaris posterior, и другую - для macula sacculi. Кроме того, от нее отходят более мелкие ветви, которые снабжают область преддверного слепого мешка улитки, нижние части utriculus и sacculus, заднюю ампуллу и eras commune с большею частью полукружных каналов. Что касается улитковой ветви этой артерии, то она входит в modiolus, снабжает нижний завиток улитки своими ветвями и анастомозирует с arteria cochlearis propria.
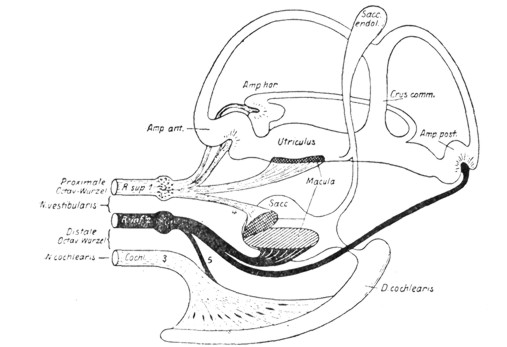 |
Рис. 24. Схема иннервации концевых аппаратов внутреннего уха кролика (по de Burlet). |
Arteria cochlearis propria входит через больший каналец tractus spiralis foraminosus вместе с нервными ветвями в modiolus и направляется к его вершине, отдавая ветви к завиткам улитки, снабжая ganglion spirale и далее разветвляясь больше по стенке scalae vestibuli.
На этом пути, кроме указанных ветвей, от arteria cochlearis propria через правильные промежутки в радиальном направлении отходят ветви, которые с одной стороны достигают эндоста верхней стенки scalae vestibuli до самой ligamentum spirale, а с другой - дают особенно богатую сеть капилляров для stria vascularis.
Кроме того, некоторые ветви направляются в lamina spiralis ossea, достигают внутренней части membranae basilaris и, проходя до туннеля Кортиева органа, образуют предкапиллярное (венозное) сплетение, которое переходит в vas spirale.
Только редкие ветви заходят за туннель для снабжения membranae basilaris, zona pectinata которой совершенно не имеет кровеносных сосудов.
Вены преддверия, сопровождая соответственные артерии, но в обратном направлении, большею частью вливаются в тонкую vena aquaeductus vestibuli, которая вначале вместе с ductus endolymphaticus, а потом в особом канальце подходит к sinus petrosus superior и впадает в него.
Меньшая часть вестибулярных вен впадает в более крупную вену - vena canaliculi cochleae, которая вблизи круглого окна вступает в довольно широкий костный канал, подходит по нему к bulbus venae jugularis, куда и впадает.
В улитке из капиллярных сетей laminae spiralis образуются вены laminae spiralis, которые вдоль ее прикрепления направляются спирально через все завитки улитки, но в основном завитке эти вены соединяются с двумя спиральными венами.
В ligamentum spirale вены собираются в области prominentia spiralis (vas prominens) и направляются по эндосту scalae tympani к оси улитки, образуя здесь сплетения; получив венозные ветви от ganglion spirale, они следуют далее к передним venae spirales (верхнего и частью среднего завитков) и к задней vena spiralis (остальной части среднего и основного завитков).
Вены ligamenti spiralis, анастомозируя с венами laminae spiralis, одновременно принимают радиальные веточки как от тимпанальной стенки laminae spiralis, так и от ganglion spirale. Наконец, все они вливаются в vena canaliculi cochleae.
Кроме того, как из вен laminae spiralis, так и из обеих спиральных вен образуются ветви, которые направляются во внутренний слуховой проход и там вливаются в vena auditiva interna, которая на своем пути принимает венозную кровь, главным образом, от n. octavus, его оболочек и от стенок meatus acusticus internus. Vena auditiva interna впадает в sinus petrosus inferior.
Лимфатических сосудов во внутреннем ухе не обнаружено. Большие перилимфатические и эндолимфатические пространства внутреннего уха имеют связь с субарахноидальным пространством, о чем уже упоминалось. Но периваскулярные и периневральные лимфатические пространства, которые можно инъицировать из субарахноидального пространства, не находятся в непосредственно прямой связи с большими лимфатическими пространствами внутреннего уха.
|
The requested URL /down.htm was not found on this server.