
The requested URL /topic.htm was not found on this server.
Известно, что в преддверии лабиринта имеются два мешечка - utriculus и sacculus - с заложенным в них отолитовым аппаратом. В состав последнего входят maculae acusticae и отолиты. Macula acustica образуется за счет опорных и чувствительных клеток, которые имеют волоски, спаянные друг с другом желатиноподобной массой. Волоски опорных клеток несколько длиннее волосков чувствительных клеток и на своей периферии, переплетаясь, образуют род войлока, называемого отолитовой мембраной. На этой мембране или, вернее, в щелях ее лежит большое число кристаллов углекислого кальция (аррагонит). Кристаллы, склеенные между собой желатиноподобной массой, образуют род камешка, называемого отолитом, или, как теперь называют, статолитом. Отолиты utriculus'ов расположены, приблизительно, в горизонтальной, отолиты же sacculus'ов лежат, приблизительно, в сагиттальной плоскости. В переднем своем отделе отолиты sacculus'ов загнуты к периферии, и эти уголки (Sacculusecke) стоят, приблизительно, во фронтальной плоскости.1 У млекопитающих (кролик) окончания вестибулярного нерва подходят к чувствительным клеткам maculae acusticae, причем дистальная ветвь нерва иннервирует большую массу maculae acusticae sacculi, проксимальная же - maculam acusticam utriculi и тот небольшой участок maculae acusticae sacculi, который относится к Sacculusecke. Эта своеобразная иннервация заставляет предполагать, что функция отолитов sacculus'ов и utriculus'ов неодинакова и, больше того, Sacculusecke, невидимому, должен отличаться в функциональном смысле от функции основной пластинки sacculus - отолита.
Раньше отолитам приписывалась слуховая функция, отчего и существует старое название macula acustica, т.е. слуховое пятнышко. Helmholtz (1870 г.), например, считал, что благодаря им мы ощущаем шумы. Breuer первый высказался в том смысле, что отолитам не свойственна слуховая функция и что, вернее всего, это есть орган статического чувства. Он же создал первую теорию об отолитовой функции. Сущность ее заключается в следующем: отолиты, как тяжелые тела, давят на волоски подлежащих чувствительных клеток, и это давление, даже при покойном положении тела, позволяет человеку определять вертикаль. Если человек совершает прямолинейное (прогрессивное) движение, то отолиты, в силу своей тяжести, по инерции будут отставать от этого движения, будут как бы скользить по macula acustica, и это скольжение, являясь раздражителем окончаний вестибулярного нерва, вызовет в мозгу представление о прогрессивном движении. Теория Breuer'a не имела фактических экспериментальных подтверждений и базировалась больше на умозрительных предположениях. Несмотря на это, все же принципиально не поколеблена она и до сих пор. Позднейшие достижения экспериментальной техники только детализировали эту теорию, но не опровергли ее.
По теории Breuer'a можно сказать, что всякая сила, изменяющая положение отолитов, должна вызывать отолитовую реакцию, и что, следовательно, специфическим раздражителем отолитового аппарата является сила тяжести их при наклонах головы, инерция (прямолинейное ускорение) в начале и в конце прямолинейных движений (в начале, например, и в конце подъема и спуска в лифте) и, наконец, прямолинейное действие центробежной силы, сопутствующее круговому перемещению (карусели, поворот при езде). Такое представление об адэкватном раздражителе отолитового аппарата за последнее время подверглось экспериментальной проверке.
>Magnus и de Kleyn, на основании добытых ими экспериментальных фактов, пришли к тому заключению, что utricul'ярные и saccul'ярные окончания вестибулярного нерва максимально приходят в раздражение в том случае, когда отолиты висят и тем самым натягивают волоски нейроэпителия. Quix высказывает совершенно противоположную точку зрения. Он считает, что максимальным раздражителем нервных окончаний является давление отолитов, при отвисании же их раздражения почти не происходит по той причине, что при отвисании отолиты будут тянуть только волоски опорных клеток, к которым они прикреплены, а волоски чувствительных клеток, не имеющие непосредственного анатомического контакта с отолитами, будут оставаться безучастными. Разногласие этих авторов сказывается еще и в другом отношении. В то время как Magnus и de Kleyn совершенно отрицают прогрессивное ускорение как раздражитель отолитового аппарата и относят его к раздражителям полукружных каналов, Quix придерживается старого взгляда Breuer'a. Наши исследования показали, что отолитовый аппарат представляется чрезвычайно лабильным в смысле своего раздражения. Оказывается, что utricul'ярные и saccul'ярные окончания вестибулярного нерва раздражаются при изменении положения тела и головы, при прямолинейном действии центробежной силы (отрыв отолитов, по Wittmaack'y) и, больше того, даже при изменении внутрилабиринтного давления. Если произвести давление на окна лабиринта, то отчетливо выступает классический отолитовый рефлекс - отклонение глаз (Хилов).
Суммируя добытые по настоящее время гипотетические и экспериментальные данные, можно сказать, что несомненными адэкватными раздражителями отолитового аппарата являются: 1) изменение положения тела в пространстве и 2) прямолинейное ускорение, развивающееся при прогрессивных перемещениях. В аспекте этих двух адэкватных раздражителей приводим две современные конкурирующие теории: 1) Magnus'a и de Kleyn'a и 2) Quix'a.
1. Рефлексы отолитового аппарата по теории Magnus'a и de Kleyn'a. Если животное (кролик) находится в нормальном положении, то отолиты utriculus'ов будут давить на волоски нейроэпителия и n. vestibularis, которые будут получать минимум раздражения. Если же животное перевести из нормального положения на угол в 180°, т.е. на спину, то отолиты utriculus'ов будут висеть, и тогда наступит максимум раздражения.
При положении животного на правом боку висеть будет только отолит правого sacculus'a и будет давать максимум раздражения, отолит же левого sacculus'a будет давить на волоски и вызывать только минимум раздражения. При положении на левом боку картина раздражений обратная. Если животное перевести из нормального положения головой вверх, то висеть будут Sacculusecken обеих сторон, и это вызовет максимум раздражения вестибулярного нерва. При тех положениях, когда наступает максимальное раздражение, появляется изменение тонуса поперечно - полосатой и гладкой мускулатуры тела, появляются тонические рефлексы, продолжающиеся до тех пор, пока удерживается приданное животному положение. Эти рефлексы называются рефлексами положения.2
Рефлексы на поперечно-полосатую мускулатуру тела (соматические). Различают следующие рефлексы положения: тонические рефлексы на мышцы конечностей, шеи и туловища, лабиринтный установочный рефлекс (Labyrinthstellreflex) и рефлексы на мышцы глаз.
Тонический рефлекс на мышцы конечностей. Если животное положить на спину, т.е. на 180° от нормального положения, то передние и задние конечности разгибаются и удерживаются в таком состоянии до тех пор, пока животное находится в приданном ему положении. Этот тонический рефлекс на разгибатели, и именно их гипертонус, зависит от раздражения utriculus - отолита, т. к. при таком положении эти отолиты висят и, следовательно, дают максимум раздражения вестибулярного нерва. Отолиты же sacculus'ов при таком положении не потягивают за волоски и не дают максимального раздражения, и в своем действии на вестибулярный нерв ничем не отличаются от действия при нормальном исходном положении. Особенно отчетливо рефлекс гипертонуса разгибателей наблюдается у децеребрированных животных. При нормальном положении животного отолиты utriculus'ов дают минимум раздражения, т. к. они тогда не висят, а давят на волоски нейроэпителия, вследствие чего тонус разгибателей ослабевает (гипотонус), но зато, по закону антагонистов (Gesetz der reciproken Innervation), сгибатели будут находиться в состоянии гипертонуса, и животное предпочитает сидеть с поджатыми, согнутыми конечностями. Если животному экстирпировать один лабиринт, то при положении максимального раздражения отолитов utriculus'ов гипертонус разгибателей всех 4-х конечностей слегка ослабевает, но не исчезает. Это говорит о том, что тонический рефлекс с отолитов utriculus'ов распространяется на конечности обеих сторон (перекрест). Если же животному удалить оба лабиринта, то гипертонуса конечностей получить уже не удается, что говорит о лабиринтном происхождении тонического рефлекса.
Тонический лабиринтный рефлекс на мышцы шеи и туловища. При положении животного на спине происходит откидывание головы, зависящее от гипертонуса разгибателей затылка, обусловленного, в свою очередь, максимальным раздражением вестибулярного нерва отолитами utriculus'ов, потягивающими волоски нейроэпителия. При нормальном положении животного, когда отолиты давят на нейроэпителий, т.е. дают минимум раздражения, тонус разгибателей готовы ослабевает, по закону же антагонизма начинает усиливаться тонус сгибателей головы, и животное удерживает ее в положении наклона вниз - вперед. Таким образом, тонический лабиринтный рефлекс на разгибатели головы аналогичен рефлексу на конечности и тоже зависит от функции utriculus - отолитов. Все же существует некоторая разница между обоими рефлексами, выступающая особенно отчетливо на животном с односторонней экстирпацией лабиринта. У такого животного, как уже сказано, остается тонический рефлекс на конечности обеих сторон, а рефлекс на мускулатуру шеи изменяется, а именно - голова животного бывает повернута в сторону оперированного уха. Таким образом, оставшийся нетронутым лабиринт посылает тонический импульс к мышцам шеи только одной стороны (перекреста нет), мышцы же шеи другой стороны, не получая импульса, остаются атоничными, вследствие чего и происходит поворот головы. Итак, utriculus-отолит стоит в функциональной связи с мышцами шеи только одной стороны, а с мышцами конечностей - обеих сторон. Тонический рефлекс на мышцы затылка исчезает, если удалить оба лабиринта, что говорит о лабиринтном происхождении этого рефлекса. В такой же зависимости от отолитового аппарата находится и мускулатура туловища.
Лабиринтный установочный, рефлекс (Labyrinthstellreflex). Под этим названием понимается такое явление, когда животное, выведенное из нормального положения тела, рефлекторно снова занимает нормальное положение. Этот рефлекс исходит из sacculus'ов. Если животное положить, например, на правый бок, то будет висеть отолит правого sacculus'a, что вызовет максимальное раздражение правого saccul'ярного нерва, левый же sacculus будет давать минимум раздражения, так как его отолит будет давить на волоски нейроэпителия. Максимум раздражения правого sacculus'a будет вызывать гипертонус определенных мышц тела, обусловливающих поворот животного в нормальное положение. Животное не может принять положения на левом боку, так как тогда произойдет максимум раздражения левого sacculus'a, возникнет гипертонус противоположной группы мышц, и животное снова должно принять нормальное положение. Равновесие наступит только в том случае, когда sacculus-отолиты обеих сторон будут расположены симметрично в отношении вертикали. Особенно демонстративно участие sacculus - отолита в установочном рефлексе проявляется на животном, у которого удален один лабиринт. Такое животное, положенное на сторону оперированного уха, не дает Labyrinthstellreflex'a, т. к. его sacculus-отолит здоровой стороны при таком положении будет давить, а не тянуть за волоски нейроэпителия, т.е. будет давать минимум-раздражения, и рефлекса не наступит. Другое дело, если положить животное на здоровую сторону, - тогда sacculus-отолит здоровой стороны будет висеть, т.е. давать максимум раздражения, и это вызовет появление Labyrinthstellreflex'a, отчего животное тотчас же примет нормальное положение. На кролике с односторонней экстирпацией лабиринта удается наблюдать Labyrinthslellreflex и без опытов с изменением положения животного. Уже в состоянии покоя кролик стремится держать голову не только приведенной к плечу оперированной стороны (рефлекс, исходящий из utriculus'a), но и поворачивает ее таким образом, что здоровое ухо бывает обращено вверх, т.е. установлено в такое положение, когда sacculus-отолит здорового уха будет находиться в состоянии минимального раздражения, т.е. когда он давит, а не висит. Если насильно повернуть голову животного в положение здоровой стороной вниз, то наступит максимум раздражения здорового sacculus'a, и животное снова повернет голову здоровым лабиринтом вверх.
Тонические лабиринтные рефлексы, на мышцы глаз. Различают два вида тонических лабиринтных рефлексов на мышцы глаз: вертикальные отклонения глаз и ротаторные движения глаз.
Вертикальные отклонения глаз. Если животное положить на один бок, например на левый, то его левый глаз окажется отведенным кверху, а правый книзу. Движение левого глаза вверх зависит от сокращения (гипертонуса) m. rect. sup. sinistri, а движение правого глаза вниз - от гипертонуса m. rect. infer, dextri. Это сокращение мышц обусловлено тем, что при положении кролика на левом боку левый sacculus - отолит висит и дает максимум раздражения нервным окончаниям, которые посылают тонический импульс к musc. rect. sup. своей стороны и к musc. rect. infer, противоположной. Правый sacculus - отолит при таком положении животного почти бездействует, т. к. он давит на волоски нейроэпителия и, следовательно, дает минимум раздражения их. Обратная картина получается в том случае, если кролика перевести в положение на правый бок. Тогда выступает действие отолита sacculus'a правой стороны, появляется гипертонус противоположных мышц, и глаза животного отклоняются в обратном направлении. Таким образом, в отношении этого рефлекса оба sacculus'a функционируют антагонистично. Их действие будет уравновешено только при таком положении кролика, когда он примет симметричное положение в отношении вертикали (напр., в нормальном положении, в положении на спине); тогда оба отолита sacculus'ов будут посылать к прямым мышцам глаз одинаковые импульсы, и никакого отклонения глаз не наступит.
Для пояснения функции отолитов sacculus'oв в отношении отклонения глаз приведем такой опыт. Удалим один лабиринт и положим животное на здоровую сторону - произойдет отвисание отолита sacculus'a здоровой стороны, т.е. максимум раздражения вестибулярного нерва, и тогда наступит вертикальное отклонение глаз. Теперь переведем животное в положение на оперированную сторону. В таком положении оставшийся sacculus - отолит будет давить, т.е. давать минимум раздражения, и никакого отклонения глаз не наступит. Если удалим и другой лабиринт, то при положении на правом и левом боку рефлекторного отклонения глаз уже не бывает, что говорит о лабиринтном характере этого рефлекса.
Ротаторное движение глаз. При положении кролика головой вверх будут висеть и потягивать волоски нейроэпителия, т.е. давать максимум раздражения, - Sacculusecke. Тогда наступит гипертонус обоих m.m. obliq. super., и глаза животного своими верхними полюсами повернутся вперед. Если же опустить кролика головой вниз, то Sacculusecke будут давить, т.е. давать минимум раздражения вестибулярного нерва, и тогда со стороны обоих m.m. obliq. super, наступит обратная реакция, а именно гипотонус; противоположная же группа мышц, т.е. m.m. obliq. infer., по закону антагонизма, будет находиться в состоянии гипертонуса, что вызовет поворот глаз верхними полюсами назад. При удалении одного лабиринта рефлекс не исчезает, но заметно ослабевает. Это говорит о том, что Sacculusecke связан функционально с m.m. obliq. sup. и m.m. obliq. infer, обеих сторон. Лабиринтный характер рефлекса доказывается тем, что после удаления обоих лабиринтов получить этот рефлекс уже не удается.
При разборе приведенных опытов с рефлексами положения может возникнуть такое возражение действительно - описанные рефлексы лабиринтного происхождения, но вряд ли можно с уверенностью сказать, что они отолитового характера, т. к. при экстирпации обоих лабиринтов разрушались не только мешечки преддверия, но и полукружные каналы, а поэтому не исключена возможность зависимости рефлексов положения от функции этих каналов. На это возражение Magnus отвечает следующим экспериментом. Он изолированно выключает отолитовый аппарат, оставляя нетронутыми полукружные каналы. Достигает этого посредством быстрого центрофугирования, когда отолиты, как тела с большим весом, по закону центробежной силы, отрываются от волосков нейроэпителия, купулы же полукружных каналов, как тела "невесомые", остаются неповрежденными. У таких животных с изолированно - выключенными отолитами все рефлексы положения исчезали, рефлексы же, исходящие из полукружных каналов, оставались неизмененными. В конце опытов лабиринт подвергался микроскопическому исследованию, которое лишний раз подтверждало, что у центрифугированных животных, потерявших тонические рефлексы положения, отолиты оказались оторванными, купулы же полукружных каналов сохранялись.
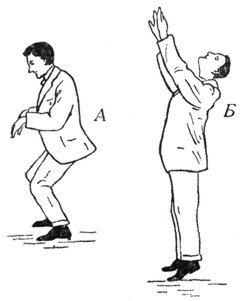 |
Рис. 7. А. Распределение мышечного тонуса конечностей и туловища при максимум - положении утрикулярных отолитов. При этом происходит сгибание конечностей и туловища. Б. То же при одном из нулевых положений тех же отолитов. Голова откинута назад под углом в 90° сравнительно с положением А. Конечности вытянуты, туловище выпрямлено (гипертонус разгибателей) (Quix). |
2. Рефлексы отолитового аппарата по теории Quix'a. При определенных положениях отолитов происходит изменение тонуса мускулатуры - тонические отолитовые рефлексы. Максимальное раздражение окончаний utricul'ярных нервов происходит в том случае, когда отолиты utriculus'ов всей своей массой давят на окончания этого нерва. Вычисления показывают, что максимальное давление отолитов происходит в том случае, если голова человека наклонена в сагиттальной плоскости вперед на угол в 30° (максимальное положение, utriculus - отолиты находятся как бы в положении "желтого пятна"). Тогда наступает максимальный тонус мышц, сгибающих тело человека в сагиттальной плоскости (сгибатели конечностей, шеи и туловища). Если же придать голове иное положение, например, наклонить еще ниже вперед или запрокинуть кверху и назад, то давление отолитов utriculus'ов будет уменьшено, и гипертонус указанной группы мышц постепенно исчезнет, а по закону антагонизма начнут функционировать противоположные группы мышц, т.е. произойдет гипертонус разгибателей конечностей и туловища (рис. 7). Благодаря действию utriculus'oB, регулирующих тонус указанных мышц, человек удерживает свое тело в состоянии равновесия. При дальнейшем изменении угла наклона наступит такое положение, когда отолиты будут висеть и, следовательно, окончания utricul'ярного нерва почти не будут получать раздражения. Тогда utriculus'ы уже не могут служить регулятором равновесия, и человек должен упасть (при таких наклонах отолиты находятся как бы в положении "слепого пятна"). При наклонах головы набок (во фронтальной плоскости) функционируют, главным образом, sacculus-отолиты. Правый sacculus усиливает тонус аддукторов левых конечностей и туловища, а левый - правых. При нормальном положении головы правый и левый sacculus'ы действуют как антагонисты, одинаково тонизируют эти группы мышц, и благодаря этому человек удерживает строго вертикальное положение во фронтальной плоскости. Если же наклонить голову на некоторый угол к правому плечу, то левый sacculus-отолит будет уже придавливать, т.е. раздражать волоски нейроэпителия, и это вызовет появление гипертонуса аддукторов правых конечностей, давление же правого sacculus - отолита при таком положении головы будет уменьшаться, что вызовет ослабление раздражения правого saccul'ярного нерва, в результате чего произойдет ослабление тонуса (гипотонус) аддукторов левых конечностей, и их место, по закону антагонизма, займет гипертонус абдукторов конечностей той же стороны. Таким образом, при наклоне головы на правое плечо произойдет аддукция правых конечностей и абдукция левых. При наклоне головы на левое плечо картина тонуса будет обратная (рис. 8). Перемена положения отолитов utriculus'ов и sacculus'ов вызывает также изменение тонуса глазных мышц, а именно utrlculus-отолиты тонизируют прямые мышцы глаз, то-есть заведуют вертикальным отклонением глаз вокруг фронтальной оси (вверх и вниз), отолиты же sacculus'ов, тонизируя косые мышцы, вызывают вращательные движения глаз ьокруг сагиттальной оси.
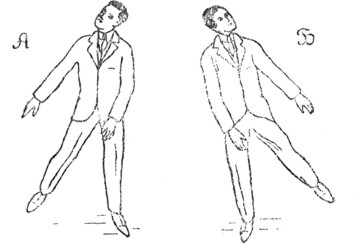 |
Рис. 8. А. Приведение левых конечностей и отведение правых при максимум-положении для правого саккулярного отолита. Это достигается таким наклоном головы при некотором повороте ее около вертикальной оси, как показывает рисунок. Б. Обратное отношение: приведение правых конечностей и отведение левых при максимум - положении для левого sacculus - отолита (Quix). |
Перечисленные группы мышц участвуют и при других раздражителях мешечков преддверия - при прогрессивных движениях и действии центробежной силы. Как пример можно привести движение на лифте (вертикальное перемещение) (Mulder). В первый момент движения наверх (прямолинейное ускорение) отолиты utriculus'ов, не успев получить приданную скорость движения, по инерции будут давить на волоски нейроэпителия, то - есть дадут максимум раздражения, в результате чего произойдет сгибание головы. При остановке движения те же отолиты будут по инерции продолжать вертикальное движение вверх, следовательно, будут отставать от волосков нейроэпителия и тем самым перестанут наносить раздражение на окончания нерва - тогда гипертонус сгибателей исчезнет, а на смену ему наступит гипертонус разгибателей, и голова поднимется кверху-кзади. Это участие отолитов utriculus'ов при прогрессивном движении особенно отчетливо наблюдается при скачке лошади (рис. 9).
Тоническое действие saccul'ярных отолитов при действии прямолинейного ускорения легко демонстрируется посредством такого эксперимента. Если голубя перемещать прямолинейно по фронтальной плоскости, например, вправо, то левое крыло в начале движения окажется приведенным к туловищу, а правое - отведенным; в конце движения - наоборот, правое приведенным, левое отведенным. Объясняется это тем, что в начале этого движения правый sacculus - отолит давит на нейроэпителий и тем самым вызывает аддукцию противоположного крыла, левый отолит, наоборот, отстает от нейроэпителия, т.е. перестает раздражать нервные окончания, вследствие чего пропадает тонус аддукторов правого крыла, абдукторы же его, потеряв своего антагониста, вызывают отведение этого крыла.
3. Рефлексы на гладкую мускулатуру (вегетативные). Вопрос о влиянии отолнтового аппарата на функцию вегетативной нервной системы находится в стадии экспериментальной и клинической разработки. Удалось установить, что при движении на четырехбрусковых качелях, когда экспериментальное животное во время качания сохраняет горизонтальное положение, появляющееся при этом, раздражение отолитового аппарата вызывает учащение и усиление дыхания. Этот вегетативный рефлекс наступает только при спуске хвостом к движению и при подъеме головой к движению, тогда как при спуске головой к движению и при подъеме хвостом к движению реакция получается обратная, т.е. ослабление и замедление дыхания. Такое влияние качания на функцию отолитового аппарата и вегетативную нервную систему объясняется тем, что при известном положении животного, последнее в противоположных пунктах качания будет испытывать периодически максимальное и минимальное состояние раздражения отолитового аппарата, т.е. наступит периодическая смена давления и отвисания отолитов (Хилов). Экспериментальное исследование отолитовых вегетативных рефлексов затрудняется тем обстоятельством, что при раздражении отолитового аппарата небезучастными остаются подвижные внутренние органы, которые при изменении положения тела животного или при движении его дают свои сенситивно - вегетативные рефлексы, наслаивающиеся на вегетативные отолитовые. Особенно резко выступают вегетативные рефлексы при раздражении отолитов во время морской и авиационной качек (Воячек, Quix). Тогда появляется вегетативный симптомокомплекс морской болезни - головокружение, тошнота, рвота, запор или понос, затруднение дыхания, появление холодного пота, бледность кожных покровов и т.д.
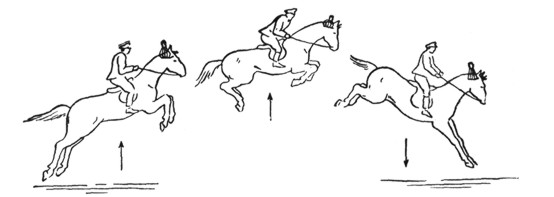 |
Рис. 9. Положение отолитов utriculus'ов и тонус мышц у лошади во время перескакивания через барьер: 1 позиция - подскок: отолит приближается к macula acustica, и давление его увеличивается ; 2 позиция - на вершине прыжка: наибольшее давление отолита, наибольшее сгибание конечностей; 3 позиция - падение: отолит отдаляется от macula acustica, давление уменьшается, передние ноги, а вслед затем и задние, выпрямляются
(разгибаются) (Quix).
|
|
The requested URL /down.htm was not found on this server.