
The requested URL /topic.htm was not found on this server.
Резонансная гипотеза Helmholtz'a. Еще в 1683 году Verney высказал предположение, что восприятие звука должно происходить в улитке по принципу созвучия - резонанса. Он предполагал, что перцепция звука происходит в lam. spiralis ossea, причем низкие тоны должны восприниматься широкой частью ее, в первом завитке улитки, высокие же звуки - в узкой части, у верхушки. С мнением Verney были согласны Valsalva, Boerhaave, Haller. В 1760 г. Cotugno, открывший лабиринтную жидкость, впервые обратил внимание не на костную часть спиральной пластинки, а на перепончатую. Предположения этих авторов были забыты, и только в 1840-1842 гг. Carus и Harnover вновь начали интересоваться вопросом о локализации звуковых вибраций во внутреннем ухе. К тому же времени относится |ряд детальных анатомических и гистологических открытий о строении внутреннего уха (Hensen, Hasse). Helmholtz первый анализировал все добытые до него данные, обработал их, математически обосновал их и экспериментально-акустически проверил. Как результат его критической работы и появилась резонансная гипотеза слуха. Для пояснения гипотезы Helmholtz'a приведем классический опыт с резонансом. Если поднять демпфер рояля и затем произнести на какой-либо высоте звук "О", то в рояле отчетливо повторится звук. Гласная "О" - сложный звук, состоящий из основного чистого тона и целого ряда обертонов. При произношении этого звука не все струны рояля резонируют на данный комплекс тонов, а только те из них, которые по высоте своей настройки будут соответствовать высоте тонов, заключающихся в произнесенной гласной, т.е. придут в колебание струны, соответствующие по высоте основному тону гласной "О" и ее обертонам. Аналогичное явление должно происходить и во внутреннем ухе. Принято считать, что основная мембрана представляет собой по длине сравнительно слабо натянутую перепонку и, наоборот, в поперечном направлении ее волоконца - струны (числом до 25000) натянуты сравнительно сильнее. Известно также, что струны основной мембраны в конечных завитках улитки значительно длиннее (по ter Kuile, в 12 - 20 раз) струн в основном завитке (рис. 1). Таким образом, волокна membr. basilaris можно рассматривать, как систему резонаторов, т.е. струн разной длины, приуроченных для созвучия на тоны разных высот. Когда стремя передает звуковую вибрацию на перилимфу, то периодическое колебание перилимфы передается и на основную мембрану, причем последняя будет отвечать на дошедшее колебание не всей своей массой, а только тем волокном, которое по своему строю соответствует периодичности колебания перилимфы. Соседние струны также, повидимому, придут в колебание и тем слабее, чем дальше они отстоят от настроенного на данный звук волоконца. Helmholtz предполагал, что топографически длинные волокна основной мембраны у верхушки улитки приходят в колебание от действия низких тонов, короткие же, у основного завитка - от высоких. Периодическое колебание волокон вызывает такое же колебание покоящихся на них частей Кортиевого органа с его волосковыми нервными клетками. Последние, вибрируя, то упираются своими волосками, то отходят от membrana tectoria, и это периодическое дотрагивание волосков нейро-эпителия, невидимому, и представляет собой механические условия для раздражения волокон кохлеарного нерва. Далее, полученные импульсы передаются центропетально в слуховые центры мозга. При таком механизме возбуждения окончаний кохлеарного нерва сохраняется признак специфичности энергии и именно каждое нервное волокно слухового нерва вместе с его периферическим рецептором возбуждается повидимому, только одним тоном, одной звуковой вибрацией определенного числа колебаний, подобно тому, как это имеет место в процессе цветоощущения. Внешний сложный звук (пример с гласной "О"), разложенный в улитке на свои компоненты, вызывая раздражение соответствующих отдельных нервных волокон, дойдя до корковых слуховых центров,- синтезируется, в результате чего человек слышит данный внешний звук в целом, со свойственной ему высотой, силой и тембром.
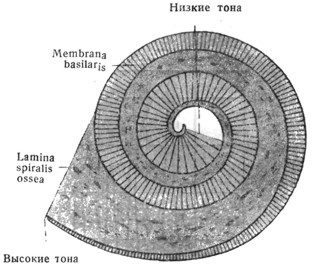 |
Рис. 1. Схематическое изображение спирального устройства улитки по Bruhl'ю, применительно к резонансной теории Helmholtz'a. |
Несмотря на ряд появившихся гипотезы о слуховой функции (Ewald, Zimmermann, ter Kuile), несогласных с мнением Helmholtz'a, все же до сих пор резонансная гипотеза должна считаться господствующей потому, что она имеет за собой экспериментальные, клинические, гисто-патологические и физические подтверждения. Munk, Baginski, Андреев, разрушая определенные части улитки собак и исследуя их слух, подтвердили правильность некоторых положений резонансной гипотезы, а именно, что глухота на дискантовые звуки всегда наступала при повреждении участков основного завитка улитки, басовая же глухота - при разрушении верхушечного. Особенно интересны и доказательны опыты Wittmaack'a, Joshii, Marx'a, v. Eicken'a, Rohr'a, Kimura и А. Попова. Эти авторы изучали влияние продолжительных, сильных звуков разных частот на внутреннее ухо. Гистологически оказалось, что высокие тоны разрушали Кортиев орган, нервные клетки и нервные волокна всегда в области основного завитка улитки, низкие же- у верхушечного. Так, например, при с5 дефект был найден только в верхней половине основного завитка улитки, остальные же части гистологически были нормальны; при h2 - выше на 0,5-1 оборот; при g - еще выше. Если раздражение производилось одновременно двумя звуками (h2 и с5), то и дефект наблюдался в двух соответствующих местах улитки (Joshii). С изучением профессиональных шумовых вредностей добыты новые сведения, подтверждающие резонансную гипотезу Helmholtz'a (Habermann, Bruhl, Zange, Kyparo, Гробштейн и др.). Лишним подтверждением положений Helmholtz'a является открытие Bezold'oM так наз. островков слуха у некоторых глухонемых. Оказывается, что патолого - анатомически у таких лиц внутреннее ухо остается сохраненным тоже островками. Наконец, известны случаи "нервной" глухоты, когда выпадает слух на высокие тоны; в таких случаях при гистологическом исследовании обнаруживается, главным образом, поражение основного завитка улитки.
Гипотезы Roaf'a - Fletcher'a, Ewald'a, ter Kuile, Meuer'a, Лазарева. Гипотеза Roaf'a - Fletcher'a представляет собой дополнение и некоторое видоизменение гипотезы Helmholtz'a. Эти авторы считают, что при воздействии звука на основную улитковую мембрану последняя приходит в колебание не отдельными своими волокнами, а по всей своей длине, но с некоторым максимумом амплитуды на определенном ее участке, имеющем определенное отношение к определенной высоте тона. Положение места наибольшей амплитуды зависит не от свойств основной мембраны, а от длины всего приходящего в движение столба лабиринтной жидкости, считая эту длину от овального окна через scala vestibtili, через место наибольшей амплитуды мембраны и далее через scala tympani до круглого окна. Известно, что масса лабиринтной жидкости в силу своей инерции представляет тем большее сопротивление прохождению колебаний,, чем выше тон. Поэтому длина столба при высоких тонах будет наименьшая, и высокие звуки замыкаются через основную мембрану вблизи ее основания, низкие же, наоборот - ближе к геликотреме. Тогда высокий звук, попадая в улитку, произведет выгиб основной перепонки с верхушкой, расположенной ближе к основному завитку улитки. Обратная картина основной мембраны будет при действии низких звуков - верхушка выгиба будет расположена ближе к геликотреме (рис. 2).
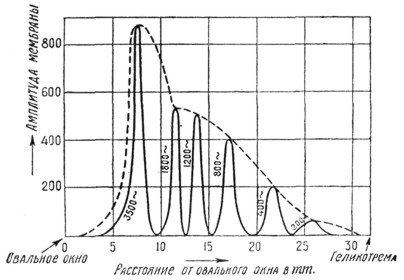 |
Рис. 2. Амплитуда основной мембраны при воздействии тонов одинаковой интенсивности на разных высотах (по Вегелю и Лэну).
|
По Fletcher'y, в мозговых центрах ощущение высоты тона получается соответственно только тому месту основной мембраны, где возникает максимальная амплитуда колебания, максимальный выгиб, независимо от того, какова будет амплитуда в других участках ее. Гипотеза Roaf'a - Fletcher'a делает понятными некоторые физические явления, трудно объяснимые гипотезой Helmholtz'a. Представим себе, что мы имеем два чистых тона в 1200 колебаний в сек. и в 500 колеб. в сек. При одновременном звучании их должны быть слышны оба звука, но на деле может оказаться, что будет слышен только звук в 1200 кол./сек. (в таком случае говорят о маскировке одного звука другим). Если начать усиливать звук в 500 кол./сек., то наступит момент, когда будут слышны оба звука. Рассмотрим в аспекте гипотезы Roaf'a - Fletcher'a, какие изменения произойдут на membr. basilaris в случае маскировки звука. Первый звук в 1200 кол./сек. производит выгиб мембраны с вершиной, стоящей ближе к геликотреме. Если теперь присоединить действие второго звука в 500 кол./сек., то окажется, что энергии этого звука будет недостаточно для того, чтобы произвести изменение формы первого выгиба, т. к. уже под его влиянием эластичность мембраны уменьшилась, и требуется уже больше усилия для образования нового заметного зубца от второго звука. Только усиливая звук в 500 кол./сек., можно получить наслоение нового выгиба на первоначальном и, следовательно, получить возможность восприятия обоих тонов.1
Ewald - наподобие оптической камеры - конструировал акустическую камеру которая в принципе напоминала строение органа слуха. Роль основной мембраны изображала узкая эластическая перепонка в 0,6 мм ширины и 8,5 мм длины. При действии звука эта перепонка колебалась, и колебания ее можно было детально рассматривать при помощи микроскопа. Ewald заметил, что при действии тонов разных высот на эластической перепонке появляются поперечные стоячие волны ("звуковые образы"), которые образуются по всей длине мембраны, причем каждый звук дает своеобразный, свойственный его высоте "звуковой образ". По мнению Ewald'a, то же самое должно происходить и на основной мембране внутреннего уха, и именно при действии звуков, подобно слабо натянутому парусу она как бы полощется и эти движения ее, разные при действии разных высот, создают разнообразные звуковые ощущения.
По ter Kuile при действии звука происходит надавливание стремени в овальное окно и это надавливание вызывает смещение перилимфы в scala vestlbuli, вследствие чего происходит выпячивание основной перепонки в сторону scala tympani; чем ниже тон, тем на большем протяжении она выпячивается и, следовательно, тем большее число концевых элементов Кортиевого органа приходит в состояние возбуждения.
По М. Meuer'y при действии звука основная мембрана вибрирует участком значительного протяжения, причем определенный участок соответствует определенному тону. Высота тона определяется числом сотрясений, испытываемых чувствительной слуховой клеткой, ощущение же силы звука зависит от числа раздражаемых клеток.
П.П. Лазарев предложил ионную теорию возбуждения, согласно которой в чувствующих клетках содержится особое вещество, которое разлагается под влиянием колебаний Кортиевых волокон. Образующиеся при этом ионы вызывают раздражение нервных окончаний Различные клетки обладают неодинаковой чувствительностью, поэтому при особых раздражениях разложение вещества происходит только в самых чувствительных клетках, при усилении раздражения захватывается все большее и большее число клеток.
|
The requested URL /down.htm was not found on this server.