
The requested URL /topic.htm was not found on this server.
Развитие ушного лабиринта у эмбриона характерно тем, что вестибулярный аппарат появляется раньше, а кохлеарный - позже; последний, как более диференцированный орган, встречается у животных, стоящих на более высокой ступени зоологической лестницы. Кроме того, у зародыша человека раньше появляется внутреннее, позже среднее и наружное ухо; филогенетически это соответствует различной степени развития каждого отдела уха у разных представителей зоологической лестницы: внутреннее, как звуковоспринимающий аппарат, в соответственной степени встречается у более низших, среднее и наружное ухо, как аппарат звукопроводящий,- у более высших представителей, у которых они функционируют как добавочная часть акустического аппарата.
И кохлеарный, и вестибулярный отделы лабиринта служат органом ориентации; механизм их ощущений состоит в восприятии вибраций: вестибулярный аппарат воспринимает более грубые, кохлеарный - более тонкие колебания. Соответственно изложенному первый обладает такими громоздкими приспособлениями, как статолиты, которые тем объемистее, чем данный представитель стоит ниже, а второй обладает весьма тонкими деталями, которые тем многочисленнее, чем данный представитель стоит выше на зоологической лестнице.
Общность роли - функция ориентации - и общность механизма восприятия вибраций обусловили, повидимому, с одной стороны, топографически общую локализацию обоих аппаратов в одном пункте (ушной лабиринт) позвоночных, с другой - дали основание для того общего названия "ухо", которое долгое время вносило путаницу в определение физиологии последнего. Отсюда происхождение недоуменных вопросов о слуховой одаренности рыб, о выборе названия для известковых образований в мешочках преддверия: отолиты или статолиты.
Современная отиатрия, уточнив функцию каждого отдела лабиринта, раскрыла также их истинную роль у разных зоологических классов: у кого более развиты элементы вестибулярного аппарата, тот обладает преимущественно развитым статическим чувством ориентации, а у кого сильнее выражены элементы кохлеарного аппарата, тот обладает преимущественно развитым акустическим чувством ориентации.
Приведенные соображения облегчают усвоение развития уха в сравнительно - анатомическом и эмбриологическом обзоре.
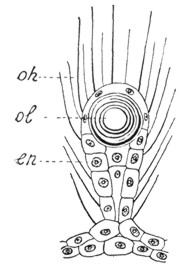 |
Рис. 1. Слуховое щупальце медузы (по Геккелю) |
У медуз имеются щупальца в виде булавовидных выростов эктодермы, содержащих круглое тяжелое образование - статолит, происходящий из энтодермы. Поверхность каждого щупальца усеяна чувствительными волосками - ресничками, передающими раздражения от колебаний окружающей среды клеткам эпителия, к которым подходят нервные волоконца (рис. 1).
Гидромедузы - Vesiculatae и Trachomedusae - обладают щупальцами, замкнутыми в виде пузырька, так называемыми статоцистами. Это - углубление эктодермы, замыкающееся, отшнуровывающееся от кожи и наполняющееся жидкостью, среди которой помещены один или несколько энтодермических камешков - статолитов; стенка такого пузырька соединена с соответственным нервом, получающим раздражения от удара статолита в стенку, как при колебании ближайшей сферы так и при перемене положения самой медузы, что вызывает перемещение статолита с определенным физиологическим последствием.
Некоторые кольчатые черви - Arenicola marina имеют эктодермические вогнутости - статокрипты, где вместо подлинных энтодермических статолитов содержатся инородные тела - песчинки.
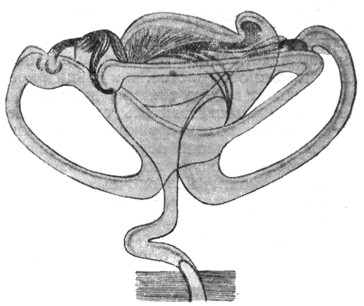 |
Рис. 2. Правый перепончатый лабиринт Acanthias vulgaris. Увеличение в 4 раза (по Retzius'y). |
То же строение наблюдается уголотурий, живущих на большой глубине. Ракообразные обладают отчетливо развитыми статокриптами в основном членике передних сяжков - antennulae, где вместо энтодермических статолитов содержатся песчинки, а вместо ресничек находятся хитиновые щетинки. У таких ракообразных, как Amphipoda, найдены замкнутые пузырьки на надглоточном узле; у мизид - на хвосте; нервы подходят из последнего узла брюшной цепи. Как и у других позвоночных, статический аппарат у рыб начинается в виде пузырька. По обеим сторонам задней части головы появляется вогнутость эктодермы; углубляясь и отшнуровываясь от кожи, она окружается мезодермическими клетками, соединяется своим дном с нервом и делится на две части: utriculus с полукружными каналами и sacculus.
Круглоротые рыбы - Myxine Glutinosa - имеют один круг, составляющий соединение двух полукружных каналов, открывающихся своими расширенными'концами - ампуллами (cristae ampullares) в общую полость, содержащую maculam statolithicam.
У селахий (рис. 2) лабиринт представляется в виде одной общей полости, в которой находятся сзади наперед utriculus, sacculus и lagena - начало улитки; полукружных каналов три, причем два из них - superior и posterior - соединяются своими свободными концами, а латеральный (горизонтальный) самостоятельным окончанием открывается в общую полость (рис. 3 и 4).
У карповидных и у сомов вестибулярный аппарат соединен с плавательным пузырем; между последним и лабиринтом лежит цепь косточек, представляющих отчленение дуг позвонков или ребер.
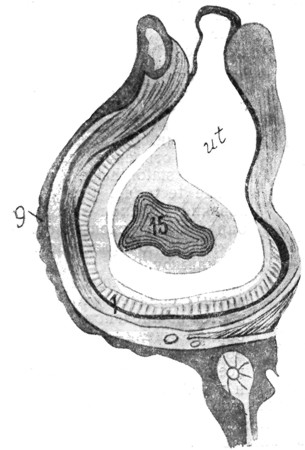 |
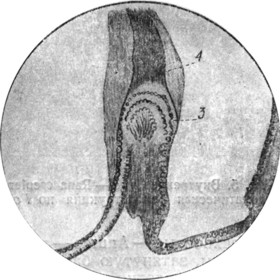 |
Рис. 3. Мешечек преддверия карася (сильное увеличение; собственный препарат). |
Рис. 4. Crista et cupula ampullaris карася (сильное увеличение; собственный препарат.) |
Рыбы не имеют улитки; это оправдывает предположение, что их лабиринт - орган чисто статический.
Однако, Frisch и его сотрудник Stetter, пользуясь методом условных рефлексов, доказали, что рыбы слух имеют: у пескаря верхняя граница определена ими в пределах от 4 до 7 тысяч колебаний в секунду; у некоторых пескарей острота слуха приближается даже к нормальному слуху человека.
Оба автора экспериментально доказали, что sacculus и lagena у рыб являются органом слуха, тогда как их utriculus и полукружные каналы служат органом равновесия (лит. 45).
Приведенные данные о слухе у рыб совпадают с наблюдениями Белоголовова (лит. 87), опубликовавшего свою работу в 1925 году, т.е. на 5 лет раньше Friscb'a.
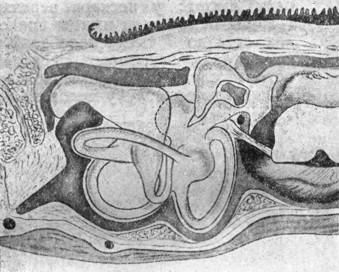 |
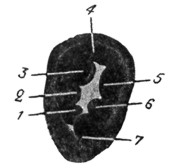
Рис. 5. Внутреннее yxo - Rana esculenta (полусхематическая реконструкция по Portmanriy). |
Сходство взглядов обоих авторов распространяется и на их оценку слуха у червей.
Белоголовов наблюдал кольчатых червей Arenicola на Белом море; Frisch наблюдал кольчатых червей Serpules в аквариуме зоологической станции в Неаполе; авторы подтверждают способность этих низших животных воспринимать звуки, не органом слуха, а тактильным чувством.
Это подтверждает высказанное выше положение, что ухо вообще есть орган восприятия вибраций; количество последних переходит в качество: грубые и менее частые вибрации воспринимаются разными формами вестибулярного аппарата; несколько менее грубые и более частые вибрации воспринимаются тактильным аппаратом; наконец, более гармонические музыкальные вибрации воспринимаются различными формами кохлеарного аппарата.
Амфибии (рис. 5), кроме мешечков преддверия и полукружных каналов, имеют уже начало среднего уха; из них Urodela имеет в слуховой капсуле овальное окно, но у нее еще отсутствует барабанная полость и Евстахиева труба. Sacculus Urodelae дает начало улитке в виде небольшого выпячивания - lagena. Utriculus, sacculus и lagena, pars basilaris которой намечает первые ступени развития органа Корти, содержатся в одной общей полости, дающей начало и полукружным каналам с их ампуллами и ампуллярными приборами. Отсюда аналогичное строение cupulae, maculae statolithicae и membranae tectoriae. Все образования перепончатого лабиринта погружены в обволакивающую их жидкость- перилимфу.
У вышестоящих позвоночных среднее ухо развивается из предгиоидной жаберной щели - единственной, удерживающейся у всех наземных, в том числе и у человека.
Бесхвостые - Anurae - имеют барабанную полость, наполненную воздухом, затянутую барабанной перепонкой и уже сообщающуюся с полостью глотки посредством Евстахиевой трубы. Между барабанной перепонкой и овальным окном отмечается косточка columella. Весь перепончатый лабиринт напоминает преддверный аппарат рыб; sacculus также имеет небольшой отросток - lagena, составляющий рудимент улитки.
У черепах Chelonidae - lagena показывает сравнительно большее развитие; у них наблюдается намек на отделение lagena от sacculus'a. Lagena рептилий вырастает еще значительнее, а у крокодилов она даже слегка изогнута, имеет развитую membrana basilaris; кохлеарный канал разделен на scala vestibuli и scala tympani; на membrana basilaris находятся нервные окончания - зачаток органа Корти. Среднее ухо рептилий имеет овальное и круглое окно, Евстахиеву трубу, columella, барабанную перепонку, лежащую на уровне кожи и у крокодилов прикрытую кожаным клапаном - зачатком наружного уха. У змей среднего уха нет; у хамелеона барабанная перепонка отсутствует.
У птиц среднее ухо ограничено костными стенками, имеет Евстахиеву трубу, columella, барабанную перепонку, лежащую в глубине короткого наружного слухового прохода, причем у совы есть клапан- зачаток ушной раковины. Внутреннее ухо птиц состоит из полукружных каналов, utriculus, sacculus и lagena с хорошо развитой raembrana basilaris. У голубя весьма мощно развиты полукружные каналы.
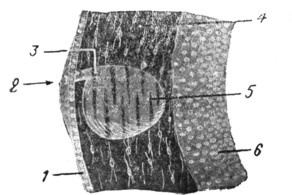 |
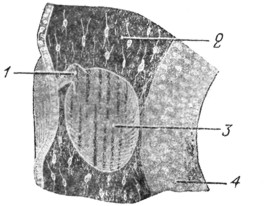 |
Рис. 6. Развитие слухового пузырька эмбриона морской свинки (по Alexander'y). Короткий каналец еще связывает пузырек с поверхностью эктодермы. |
Рис 7. Короткий каналец превращается в эндолимфатический канал; пузырек увеличился в размере. Обозначения те же, что и на рис. 6. |
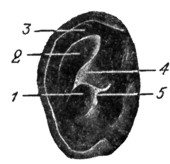
У млекопитающих (рис. 6, 7, 8, 9, 10 и 11) уже сильнее развита lagena, принимающая спиральную форму и получающая впервые название улитки. У сумчатых и одноутробных улитка изогнута слабовато; у кита уже есть 1,5 завитка; у человека - 2,5 завитка; у кошки - 3; у свиньи 4; у грызунов - 5 завитков. Орган Корти у всех млекопитающих высоко диференцирован; в среднем ухе отмечаются три слуховые косточки: молоточек, наковальня и стремячко; Евстахиева труба отлично развита; наружный слуховой проход и ушная раковина вполне закончены - кроме некоторых морских и подземных представителей, не нуждающихся в резонаторном аппарате. Ушная раковина состоит из хряща, движется с помощью мускулатуры, атавистически сохраняющейся у приматов, раковина которых, как правило, неподвижна.
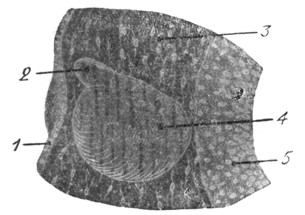 |
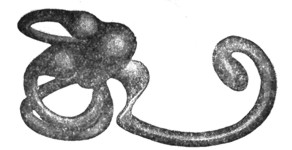 |
Рис. 8. Эндолимфатический канал уже отделился от эктодермы; слуховой пузырек еще больше увеличился в размере. Обозначения те же, что и на рис. 6. |
Рис. 9. Правый перепончатый лабиринт Echidna aculeata (по Alexander'y). |
Чувствительные клетки, происходящие от эктодермы, как известно, диференцируются в специальные органы.
Клетки эти приходят в связь с корою головного мозга посредством ганглиев, обычно имеющих два отростка: один - периферический, другой - центральный.
Положение ганглия в разных органах различно: в глазу, например, он включен в воспринимающий слой чувствительных клеток сетчатки, в ушном лабиринте он расположен вне органа Корти, вне статолитовых пятнышек и вне crista ampullaris.
 |
Рис. 10. Левый перепончатый лабиринт морской свинки. |
У рыб между продолговатым и головным мозгом существует regio acustica lateralis, где оканчивается вестибулярный нерв и "боковая линия"; это regio рассматривается, как зачаток мозжечка; он функционально связан с вестибулярным аппаратом.
У млекопитающих видно, как вестибулярный аппарат связан с мозжечком волокнами, идущими к nucleus tecti, отчасти к nucleus dentatus; отсюда нейроны направляются к красным ядрам через ножки мозжечка и затем в tractus rubro-spinalis и к коре головного мозга.
Как вестибулярный аппарат является главным органом, так и мозжечок является главным ганглием, проприоцептивной, воспринимающей положение тела в пространстве, системой всего тела: к мозжечку приходят волокна нервов от мышц, сухожилий, суставов, оканчиваясь в nucleus tecti, от которого, как уже сказано, идут волокна к коре головного мозга.
Этим осуществляется рефлекс автоматического управления положением тела в пространстве.
Прямой, непосредственной связи между вестибулярным аппаратом и головным мозгом не существует ни у одного из зоологических классов.
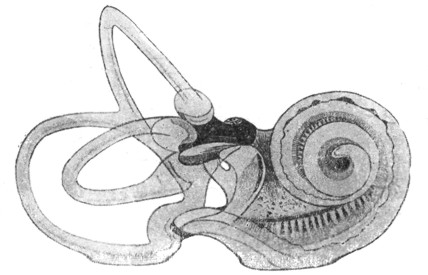 |
Рис. 11. Перепончатый лабиринт человека (по Retzius'y). |
Филогенетически мозжечок существует только у тех зоологических видов, у которых хорошо развит автоматизм; самостоятельных инициативных импульсов мозжечок не проявляет.
Исследования автора (лит. 49) убедили его в том, что:
Такое различие формы и веса статолитового аппарата указывает, что выводы, получаемые при экспериментах на лабиринте животного, могут быть полностью отнесены лишь к соответственному виду последнего, но не могут переноситься целиком и полностью на другие, хотя бы и близкие ему, виды животных.
Вестибулярный период филогенеза переходит в кохлеарный: до появления коры мозга нет и улитки.
Появление улитки совпадает с началом развития коры мозга: улитка поэтому относится невропатологами к дискриминативным органам восприятия, т.е. воспринимающим более тонкие и частые вибрации.
Кохлеарный нерв развивается позже; он присоединяется к нерву вестибулярному и таким образом создает смесь нервов звукового и статического аппаратов.
И такое же смешение звуковых и статических нервов отмечается неврологами в ядерных окончаниях.
То, что раньше называли stria acustica, теперь называют stria medullaris, так как она ничего общего со слухом не имеет.
Различие между кохлеарным и вестибулярным нервами - в их рецепторах, в их окончаниях. В остальном они перемешаны.
Когда животное переходит к наземному образу жизни, лишь тогда начинает развиваться кохлеарный аппарат. Аствацатуров говорит, что кохлеарная функция влияет на статический аппарат: когда играют гамму от басов к дискантам, животное поднимается, когда же гамма нисходит с верхних регистров до нижнего, животное опускается.
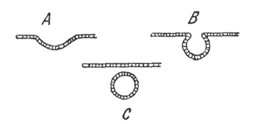 |
Рис. 12. Схема развития слухового пузырька (по Холодковскому) |
Своеобразное устройство слухового аппарата имеют насекомые. У них различают хордотональные и тимпанальные слуховые аппараты. Первые, т.е. хордотональные аппараты, состоят из нерва, нервных клеток и так называемой слуховой палочки; все это вместе расположено под хитиновым слоем кожи; когда звуковые волны вызывают сотрясение - вибрирование хитинового слоя, получается раздражение слуховой палочки, от которой ближайшие нервные клетки передают раздражение к соответственному нерву. Вторые, т.е. тимпанальные аппараты, найдены у сверчка, саранчи, кузнечиков и т.п. насекомых, издающих резкое стрекотание. Их тимпанальный слуховой аппарат состоит из перепонки, расположенной на конечностях насекомого. Иногда перепонка защищена складкой, как бы наружным ухом. Под перепонкой находится слуховой валик, состоящий из палочек и чувствительных клеточек; этот орган служит для восприятия звуков стрекотания.
Венский ученый J. Regen (лит. 45) на серии экспериментов доказал, что насекомые действительно слышат; что их стрекотание передается по воздуху, т.е., что и орган их слуха воспринимает звуки, передающиеся по воздуху; наконец, что тимпанальный аппарат сверчка, саранчи, кузнечика и т.п. насекомых является органом, воспринимающим звуки, т.е. является несомненным органом слуха. Первый зачаток внутреннего уха человека (рис. 12, 13) наблюдается у 4-недельного зародыша в форме углубления эктодермы по обеим сторонам красного мозга. Углубление это в виде пятнышка расширяется, постепенно становится шарообразным и только узким канальцем открывается наружу. Удлиняясь и суживаясь, каналец закупоривается у своего наружного отверстия, замыкаясь эпидермисом, оставляя на короткий период небольшой след клеток от своего бывшего отверстия.
В этой стадии слуховой пузырек состоит из saccus communis и ductus endolymphaticus (рис. 14).
В дальнейшем происходит диференциация верхнего и нижнего отделов слухового пузырька: из первого образуются начатки полукружных каналов, из второго - начаток канала улитки.
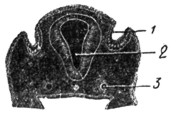 |
Рис. 13. Эмбрион человека 12 дней. Вогнутость эктодермы указывает место развития будущего лабиринта. |
Возникшие полукружные каналы постепенно удлиняются, принимают направление будущего сагиттального, фронтального и горизонтального полукружных каналов, причем одновременно углубляется диференциация верхнего отдела от нижнего, а начаток улитки, удлиняясь все больше, изгибается спирально.
Следующий период является завершением развития заднего отдела лабиринта: фронтальный и сагиттальный каналы обнаруживают свое общее колено, изогнутые части их получают угол в 90° по отношению друг к другу; их внутренние полости принимают овальную форму, а свободные концы ампуллярно расширяются, вступая в sinus utricularis superior. Позже упомянутых развивается горизонтальный полукружный канал. К этому времени полностью развит уже и ductus cochlearis.
Постепенно отчетливее вырисовывается sinus utricularis anterior - recessus utriculi; ампуллы четко отграничиваются от своих полукружных каналов; оформляется saccus endolymphaticus, sinus utricularis inferior и ductus reuniens, a sacculus полностью отделяется от utriculus'a, составляя характерное отличие лабиринта млекопитающих от других позвоночных.
В многослойном эпителии, образующем в ранней стадии эмбриона человека стенки его перепончатого лабиринта, встречаются скопления цилиндрических клеток, резко отличающихся от остального покрова слуховой бляшки.
 |
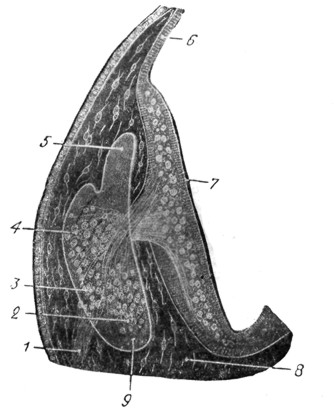 |
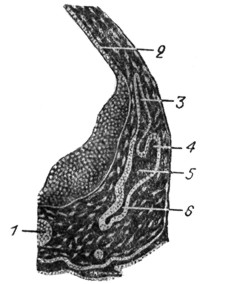
Рис. 14. Эмбрион человека пяти недель. Срез левого слухового пузырька. Вверху намечен уже эндолимфатический канал; на противоположном конце-кохлеарный канал. Пузырек заполнен эндолимфой. Вся медиальная стенка занята многослойным невроэпителием. Здесь берут начало sacculus и utriculus. |
Рис. 15. Эмбрион человека 3,5 недель. Слуховой пузырек сильно увеличен (по His'y) Срез через ganglion acusticofaclale в слуховом пузырьке. Видно близкое соседство ганглиозных клеток зачатка будущего слухового нерва с его ganglion vestibulare и ganglion cochleare (впоследствии ganglion spirale) и между ними зачаток лицевого нерва с его ganglion geniculi. |
Эти скопления постепенно разрастаются, главным образом в направлении окончания нервных волокон, идущих от ganglion octavi; здесь и надо искать первые очаги нервных окончаний внутреннего уха, носящие пока название macula communis (рис. 15).
Рост и диференциация maculae communis в сторону улитки и преддверия происходит параллельно с развитием всего лабиринта. На первых порах зачатки улитки и преддверия остаются соединенными общим невроэпителиальным мостиком; позже данный участок рассекается эпителиальной стенкой; до возникновения диференцированных полукружных каналов отмечается во внутреннем ухе эмбриона два невроэпителиальных пятнышка: одно для верхнего отдела лабиринта, другое - для его нижнего отдела (рис. 16).
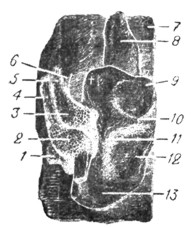 |
Рис. 16. Эмбрион человека пяти недель. 10,2 мм длины. Левый слуховой пузырек. Непосредственно к нему прилегают ganglion vestibulare и cochleare, между ними ganglion geniculi лицевого нерва (по His'y). |
С появлением ампулл от верхнего отдела невроэпителиального пятна отделяются cristae ampullares фронтального и горизонтального полукружных каналов, остатки же этого пятна служат для образования maculae utriculi. От этих клеток образуются в ампуллах зачатки cupulae ampullaris, а в macula utriculi - зачаток статолитовой перепонки.
Указанные явления происходят у эмбриона на четвертом месяце его развития.
Онтогенетически cupula и статолитовая перепонка происходят из опорных или из длинных волосковых клеток.
Статолиты, состоящие из особого вещества- арагонита - происходят от чувствительных клеток.
И cupulae, и статолитовые перепонки человеческого эмбриона впервые намечаются в виде студенистой эпителиальной массы, когда зародыш имеет около 25 мм длины, и остаются без диференциации до 125 мм роста эмбриона. Начиная же с последнего размера различают первые их очертания.
Невроэпителий нижнего отдела служит для образования: ампуллы с ее crista ampullaris и cupula сагиттального канала (рис. 17), maculae saccularis и, наконец, papillae basilaris cochleae будущего органа Корти.
Из гомогенного верхнего слоя maculae мешочков выделяется статолитовая перепонка. Статолиты в обоих мешочках преддверия появляются одновременно (рис. 18).
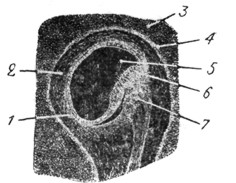 |
Рис. 17. Эмбрион человека трех месяцев. Разрез ампуллы. Плоский эпителий выстилает внутреннюю стенку ампуллы; только в средине стенки выдается бугорок с чувствующим эпителием, с приближающимся к нему нервом. Снаружи перепончатая ампулла окружена студенистой тканью, изнутри заполнена эндолимфой. |
Не трудно понять, что неравномерное развитие отдельных участков невроэпителия может привести к различным вариантам лабиринта у людей нормальных - с преобладанием чувствительности статической функции у одних, кохлеарной функции - у других, но может привести и к врожденным аномалиям в форме полного отсутствия чувствительности кохлеарного аппарата у одних или вестибулярной функции у других. В этом причина некоторых аномалий и уродств, как то: врожденная глухота, врожденное выпадение способности воспринимать звуки определенных колебаний, врожденная невыносливость к некоторым видам перемещения тела в пространстве и т.д. Первый зачаток эндолимфатического мешка найден у эмбриона восьми недель. Около этого периода sacculus отделяется от канала улитки посредством ductus reunlens, сужение которого наступает позже. Канал улитки в это время составляет как бы ответвление преддверия.
Последняя стадия эмбрионального развития внутреннего уха характеризуется возникновением canalis utriculo-saccularis, воронкообразным расширением sacculus'a и общим увеличением объема преддверия. Полости же перепончатого лабиринта суживаются, резче отделяясь от окружающих их тканей. Все отделы лабиринта в указанный период еще сообщаются между собою, и только в конце второго или в начале третьего месяца жизни эмбриона происходит полное отделение sacculus'a.
Уже в первые недели развития эмбриона человека на участке, где слуховая бляшка входит в контакт с периферическими зачатками слухового нерва, намечается многослойный цилиндрический эпителий; вскоре вокруг последнего образуются кутикулярные скопления, топографически соответствующие тельцам, дающим позже начало окончаниям нерва в cupulae et cristae ampullares в статолитовой перепонке, в подушке maculae statoliticae и в membrana tectoria органа Корти.
Эмбрион человека на пятом месяце имеет уже и cupulae, и cristae ampullares, тогда как maculae в мешечках преддверия еще покрыты только многослойным эпителием - вместо статолитов. Мезенхима в этом возрасте заполняет перилимфатические пространства внутреннего уха. В процессе обратного развития мезенхимы в перепончатом кохлеарном канале образуется membrana basilaris.
На этой базальной стенке возникают эпителиальные валики, один больший, другой меньший, образующие впоследствии эпителиальные клетки для sulcus spiralis internus и limbus spiralis, с одной стороны, и волосковые и опорные клетки, а также клетки Hensen'a - с другой.
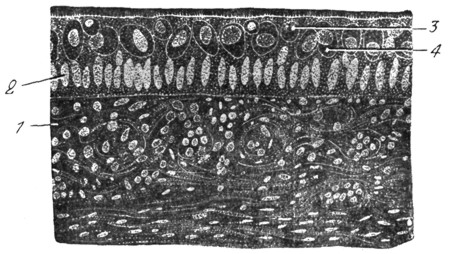 |
Рис. 18. Эмбрион морской свинки. Эпителий sacculus'a. Видны волосатые клетки, опорные клетки и соединительно - тканный слой под ними (по Alexander'y). |
С появлением внутренних волосковых клеток в большем эпителиальном валике начинает увеличиваться и crista spiralis и выступает отчетливее limbus spiralis, достигающие полного развития лишь с появлением sulcus spiralis internus.
Из покровного слоя этих двух эпителиальных валиков развивается membrana tectoria, сперва в форме кутикулярной перепонки общего ductus cochlearis.
От основания улитки до ее вершины отмечается контакт между membrana tectoria и волосковыми клетками.
В случае врожденной аномалии отмечается отсутствие этого контакта между membrana tectoria и волосковыми клетками, иногда даже с более или менее развитой степенью отрыва этой перепонки от органа Корти.
С появлением и развитием scala vestibuli происходит параллельное формирование и утончение membranae basilaris, превращающейся на своем периферическом краю в двуслойный ряд эпителиальных клеток, происходящих с эндолимфатической стороны от эпителия внутреннего уха, а со стороны перилимфатической - от мезенхимы scalae vestibuli.
Из мезенхиматозных и эпителиальных клеток берут начало stria vascularis и ligamentum spirale.1
В разных завитках улитки эмбриона в разной степени развивается papilla basilaris: у основания улитки она развита больше, чем у ее вершины. Как правило, следует отметить, что развитие улитки у эмбриона человека происходит постепенно, начинаясь у основания и заканчиваясь у ее вершины. Поэтому в различных стадиях у эмбриона человека можно наблюдать несколько видов papillae basilaris; она развита полнее и лучше у основания улитки и слабее у ее последнего завитка.
В нижнем конце базального завитка пятимесячного эмбриона человека уже почти полностью развиты волосковые клетки. Орган Корти большею частью значительно диференцирован. У восьмимесячного эмбриона уже видны клетки Hensen'a, и только membrana tectoria в некоторых участках вершины улитки показывает еще свою эмбриональную связь с поверхностным слоем диференцированных наружных волосковых клеток; но у основания улитки, в ее первом завитке у того же восьмимесячного эмбриона этих остатков уже не видно.
На четвертой неделе у эмбриона человека ganglion octavum лежит на стенке ушной бляшки, вместе с которою делится на верхний отдел и нижний. Впоследствии разветвления от верхнего отдела иннервируют macula utriculi, - cristae ampullares фронтального и горизонтального полукружных каналов; разветвления от нижнего отдела дают волокна для macula sacculi и ампуллы сагиттального канала, и для ganglion spirale улиткового нерва. Последний т.е. ganglion spirale, заметен у 10-мм эмбриона человека.
От нижнего отдела отходят волокна к nervus intermedius.
Кохлеарный нерв развивается позже вестибулярного; это говорит в пользу установившегося взгляда, что улитка позже развивается сравнительно с преддверием.
В том же эмбриональном периоде намечается путь n. facialis с отходящею от него chorda tympani.
Канал лицевого нерва составляется из трех отделов: часть от внутреннего слухового прохода до первого колена происходит из chondrocranium'a, средняя часть - из стенки лабиринта, обволакивающей во время окостенения эту часть нерва, захватывая одновременно и muscul. stapedius хрящевым кольцом, который позже оссифицируется параллельно каналу лицевого нерва, образуя с ним нечто вроде двустволки.
Нижний отдел канала n. VII образуется из височной кости, в будущем foramen stylomastoideum.
Иногда средняя часть канала VII пары новорожденного имеет дегисценцию, никогда не зарастающую. Отсюда можно понять те крайне редкие случаи паралича лицевого нерва, который совпадает с острым простудным мезотимпанитом: близость обнаженного ствола нерва делает его восприимчивым к простудным факторам.
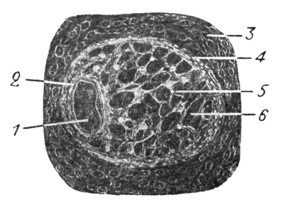 |
Рис. 19. Эмбрион человека около трех месяцев. Срез перепончатого полукружного канала. Видна тонкая стенка; в полости эндолимфа, снаружи студенистая ткань в виде сетки среди перилимфы. Наружная капсула вся из хряща и надхрящницы. |
Врожденная асимметрия лица может найти объяснение тоже в конституциональном варианте лицевого нерва, например, в гипоплязии одной его половины или отдельных веток одной его стороны, проходящих через различные участки уха.
Перилимфатические пространства образуются путем резорбции мезодермальной ткани вокруг перепончатого лабиринта; в местах неполной резорбции остаются тяжи, напр., вокруг ампулл, возле медиальной стороны utriculi и на периферической стороне перепончатых полукружных каналов. Тот же процесс латерально от стенок мешечков преддверия ведет к образованию cisternae perilymphaticae vestibuli. Одновременно с этой перилимфатической областью совершается развитие scalae vestibuli и зачаток membranae basilaris улитки, а дальнейшая резорбция мезодермальной ткани внутри последней заканчивает развитие scalae tympani и scalae vestibuli, причем последняя развивается несколько позже, чем первая. Развитие обеих протекает по направлению от основания к конечному завитку улитки. Параллельно с ними развивается и aquaeductus cochleae.
К концу полного созревания зародыша мезодермальная ткань(рис.19) полностью исчезает в полукружных каналах - сперва в сагиттальном, затем во фронтальном и в горизонтальном каналах, наконец в улитке и в aquaeductus cochleae. В пятом месяце утробной жизни перилимфатические и эндолимфатические пространства лабиринта окончательно оформлены.
В конце пятого месяца впервые в мезодерме эмбриона появляются вокруг перепончатого лабиринта хрящевые полосы; в различных направлениях от них вырастают сосуды, по которым впоследствии идет процесс окостенения: первый пункт последнего отмечен на улитке вблизи преддверия, следующий пункт - вблизи ампулл фронтального и горизонтального полукружных каналов; третья точка окостенения появляется снаружи от тела улитки в области промонториума; наконец, последние точки отмечены медиальнее будущей fossae subarcuatae, сагиттального полукружного канала и несколько впереди последнего.
В конце 4-й недели эмбриональной жизни человека из боковой и дорзальной частей его первой жаберной щели появляется несколько бугорков и валиков: два впереди, один позади и три еще более кзади. Первые три бугорка - мандибулярные, последние три - гиоидальные. В этом периоде указанные образования еще не свернуты в трубку.
К началу пятой недели (рис. 20, 21,22) возникают пять горизонтальных складок между будущим helix ascendens и нижним краем ушного отверстия, составляя сравнительно-анатомический рудимент длинных складок ушной раковины длинноухих животных. Под эпителием первой ушной складки у трехмесячного эмбриона впервые намечается ушной хрящ в виде бляшек. Хрящевая бляшка козелка имеет связь с хрящевой бляшкой противокозелка, но еще не связана с хрящом helix'a. У новорожденного хрящевые части полностью связаны своими задними краями, передние же края связаны неполно.
 |
 |
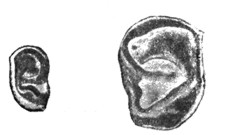 |
Рис. 20. Эмбрион человека пяти недель. Развитие ушной раковины. Видны аурикулярные бугорки будущих: завитка, противозавитка, козелка и противокозелка. |
Рис. 21. Эмбрион человека 8 недель. Ясно очерчены завиток, противозавиток, бугорок козелка и противокозелок. |
Рис. 22. Слева ушная раковина эмбриона человека шести месяцев в натуральную величину. Справа - ушная раковина новорожденного мальчика; на завитке резко обозначен бугорок Дарвина (по Schwalbe). |
Слияние хрящевых частей в замкнутую систему совпадает с окончательным оформлением наружного слухового прохода; incisurae Santorim указывают на бывшее раньше деление хряща на три части. Длина наружного уха у четырехмесячного эмбриона человека равна 7-10 мм; увеличиваясь ежемесячно на 4 мм, длина эта к концу беременности достигает 30 мм, к концу первого года жизни - 48 мм, к концу третьего года - 53 мм, в 15 лет бывает около 7 см; на этом размере рост останавливается, соответствуя среднему варианту наружного уха взрослого человека. Ушная раковина к старости удлиняется.
У птиц встречается зачаток наружного уха в виде короткого наружного слухового прохода; у совы находим клапан, дающий намек на ушную раковину.
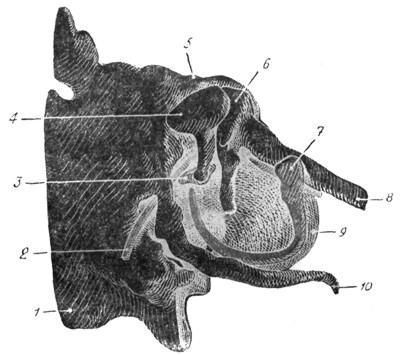 |
Рис. 23. Эмбрион человека 8 см длины. Барабанная полость (сильно увеличена). Annulus tympanicus очерчивает границы отсутствующей барабанной перепонки. Молоточек связан с хрящом Meckel'я. Небольшой бугорок указывает место будущего сосцевидного отростка. |
У млекопитающих, кроме китов, тюленей, крота, ящерицы и некоторых других, наружное ухо развито вполне и при этом состоит также из хрящей, а мускулатура, делающая подвижной ушную раковину, будучи сильно развитой у одних, остается еще у тех, у кого раковина обычно неподвижна (приматы).
Дарвинов бугорок на краю helicis соответствует задне-верхнему углу зачатка уха 4 - месячного эмбриона; сильно развитый у обезьяны, он бывает совершенно незаметным у человека. Однако встречаются ушные раковины у людей с недоразвитой ушной мочкой и весьма развитым бугорком Дарвина, а также с большой мощностью ушных мышц, приближающих такое ухо к типу соответственных видов. Варианты эти относятся к области антропологии, так как некоторые национальности имеют свои особенности в данном отношении, и к области уродств, так как недоразвитость этих отделов или, напротив, чрезмерное развитие их превращает количестненное нарушение тканевых элементов в качественную диспропорцию с соответственной асимметрией и дисгармонией, являющейся уродством в общепринятом смысле.
Развитие среднего уха представлено на рис. 23, 24, 25, 26.
Зачатки слуховых косточек находятся в бластеме первого и второго висцерального листов: из первого развивается молоточек и наковальня, из второго - стремячко.
Зачаток стремени обнаружен у зародыша на 4-й неделе его развития; зачаток молоточка и наковальни - у эмбриона 6 недель. Молоточек и наковальня происходят от первой жаберной дуги, а стремячко и шиловидный отросток - от второй жаберной щели.
До конца второго месяца слуховые косточки и капсула лабиринта человеческого эмбриона еще не оссифицированы.
В этом периоде происходит образование овального и круглого окон и кольца подошвы стремячка. Muscul. tensor tympani намечается в конце восьмой недели; muscul. stapedius - в конце 12-й недели.
Генетически оба эти мускула принадлежат к одной группе с muscul. stylo-hyoideus и muscul. biventer.
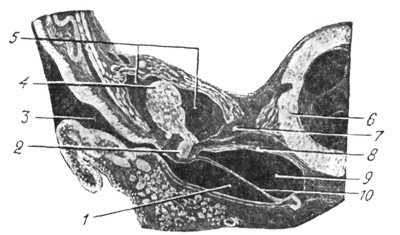 |
Рис. 24. Эмбрион человека четырех месяцев; 11см. длины. Фронтальный разрез через барабанную полость. Захвачены: наружный слуховой проход, в поперечном разрезе барабанная перепонка, часть молоточка с коротким отростком, muse, tensor tympani и капсула улитки. В полости видна эмбриональная студенистая ткань (сильно увеличено) (по Hammar'у) |
Одновременно очерчивается ostium Евстахиевой трубы; хрящ, последней встречается в виде зачатка у эмбриона человека в возрасте 4-х месяцев; большая же часть трубы в этом периоде образуется из мезодермальной ткани; ее фарингеальная часть имеет уже цилиндрический эпителий. Евстахиева труба происходит от первой жаберной щели. У эмбриона она состоит главным образом из бесструктурной массы; к моменту родов отмечаются хрящевые клетки; с возрастом этот хрящ уже не сплошной, а представляется в виде островков, и часть его оссифицируется. У различных людей имеются большие индивидуальные варианты как в отношении длины костной и хрящевой половины трубы, так и формы и диаметра ее просвета и положения обоих ее отверстий.
Детская труба короче, эластичнее и прямее; труба у взрослого более изогнута, упруга и длиннее.
Глоточное отверстие трубы у эмбриона лежит ниже горизонтальной плоскости твердого неба; у новорожденного - достигает уровня этой плоскости; у четырехлетнего ребенка оно стоит уже на 3 мм выше последней; у взрослого - на 10 мм выше.
И наружный вид, и форма, и положение глоточного отверстия Евстахиевой трубы у людей, по Урбанчичу, представляют значительные варианты даже независимо от возраста человека. То же самое можно сказать и в отношении ее тимпанального отверстия.
Оссификация головки молоточка и тела наковальни начинается в четвертом месяце, продолжается в пятом месяце параллельно с центральной частью подошвы стремячка.
Слуховые косточки видимы уже у трехмесячного эмбриона, хотя без резко очерченных контуров. Их окостенение начинается в конце третьего - в начале четвертого месяца: сперва одна только головка молоточка имеет тонкую костную оболочку; в стремени одно ребро показывает уже точку начала окостенения.
На шестом и седьмом месяцах стремя большею частью оссифицируется. Остальные части слуховых косточек еще остаются хрящевыми до конца седьмого и начала восьмого месяца, когда оссификация их заканчивается.
На девятом месяце эмбриональной жизни слуховые косточки молоточка окостеневают, а рукоятка молоточка и подошва стремени хранят остатки хряща.
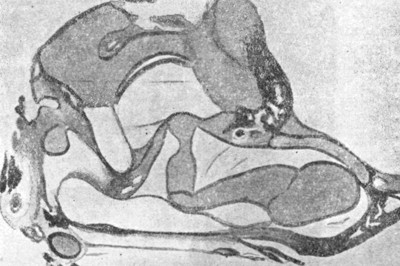 |
Рис. 25. Фронтальный разрез среднего уха эмбриона человека пяти месяцев (по Соловцову). |
Существуют значительные варианты в этом смысле: у одних слуховые косточки абсолютно оссифицированы и состоят из компактной кости с многочисленными Гаверсовыми каналами; у других неполностью оссифицированы, и в косточках находят центральное мозговое вещество.
К моменту рождения все слуховые косточки достигают своих окончательных размеров.
У новорожденных барабанная перепонка лежит почти горизонтально. Наружный ее эпидермис развит чрезвычайно сильно, разрыхлен околоплодной жидкостью.
В слуховом проходе находятся и отслоенный эпидермис и vernix caseosa.
 |
Рис. 26. Передне - задний разрез среднего уха эмбриона человека восьми месяцев (по Соловцову). |
Поэтому новорожденный в первые дни не слышит ничего; он не реагирует даже на самые сильные дисгармонические звуки, точно так же, как и котята, и щенки, которые в первые дни жизни имеют еще кожный покров на раковине, разрывающийся лишь постепенно. Постепенно наружный слуховой проход очищается, барабанная перепонка медленно принимает вертикальную форму и свой глянцевитый оттенок. Этот процесс очищения барабанной перепонки в редких случаях развивается несколько больше обычного, что может повести к развитию эпидермоидных жемчужин или даже к холестеатоме.
Когда общий зародышевый участок слухового прохода и Евстахиевой трубы уже намечен, барабанной перепонки еще нет. Лишь позже создается нижний, а затем и верхний ее отделы; верхний отдел это - будущая тонкая membrana flaccida Schrapnellii, лишенная характерных волокон -нижней части membranae tympani, т.е. substantia propria. У некоторых наблюдается неполное срастание этих двух отделов барабанной перепонки; отсюда редкие случаи врожденного отверстия или канала в барабанной перепонке - последних остатков первой висцеральной щели.
Рост барабанной перепонки можно определить по размерам annulus tympanicus эмбрионов.
Troltsch, исследуя размеры (ширину и высоту) барабанной перепонки эмбриона человека, нашел следующее:
| Возраст эмбриона | Размеры барабанной перепонки (в мм) | |
| высота | ширина | |
| 11 недель | 2 | 1,25 |
| 14 недель | 3 | 2 |
| 16 недель | 4,5 | 2 |
| 20 недель | 7 | 5,5 |
| 22 недель | 8 | 7 |
| 24 недель | 8,5 | 8 |
| 7 месяцев | 9 | 8 |
| 9 месяцев | 9,75 | 8,5 |
Annulus tympanicus, в которое вставлена барабанная перепонка, у зародыша является одною из трех частей, постепенно срастающихся и образующих всю височную кость. А именно: pars squamosa, или чешуя, pars petrosa, или пирамида, из боковой части которой впоследствии образуется сосцевидный отросток, и pars tympanica в виде одного костного ободка, annulus tympanicus, дающего затем развитие костной части наружного слухового прохода.
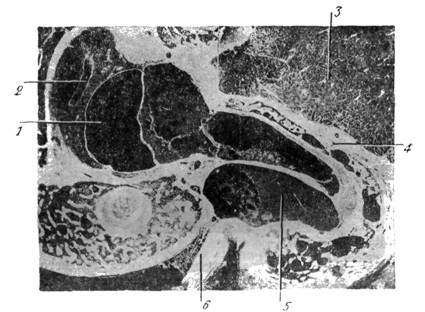 |
Рис. 27. Вертикальный срез уха на уровне utriculus (1), sacculus (2) и promontorium (4). Эмбрион человека пяти месяцев (по Alexander'у). |
У эмбриона annulus tympanicus с барабанной перепонкой лежит горизонтально, так что последняя составляет часть основания черепа. У новорожденного это положение еще сохраняется. Только постепенно - с дальнейшим ростом ребенка и развитием черепа - барабанная перепонка выпрямляется.
У восьмимесячного эмбриона суставы и связки слуховых косточек уже вполне развиты. Барабанная полость заполнена миксоидной тканью. Однако на уровне слуховых косточек уже появляются свободные пространства („карманы") микроскопической величины.
Начало средней трети барабанной полости отмечается у эмбриона одного месяца; передняя ее треть развивается на шестой неделе, после чего труба, начинающаяся в глотке, своим длинником внедряется в барабанную полость. Барабанная перепонка развивается также сперва в своей передней части и лишь к концу образования полости среднего уха развиваются ее задние отделы. В общем различают три периода в развитии среднего уха: 1-й период - до 7 недель, соответствует первичному образованию барабанной полости, 2-й период - до 3 месяцев, соответствует образованию трубно - барабанного канала, 3-й период, являясь завершением развития среднего уха, продолжается еще и в постэмбриональной жизни (рис. 27). Во втором периоде впервые отмечается в среднем ухе миксоидная ткань, достигающая полного развития у зародыша 6-7 месяцев; ткань эта расположена под слизистой оболочкой и способствует внутризародышевому оформлению среднего уха. В целом височная кость эмбриона составляется из следующих частей: коробка среднего уха из начатков кости покрышки; наковальня и молоточек - из первой, а стремячко и шиловидный отросток - из второй жаберной дуги. По Гёксли, petrosum составляется из трех окостеневающих частей: prooticum, opisthoticum, epioticura.
Слизистая барабанной полости к концу девятого месяца тоже занята еще миксоидной тканью, но уже начинают вырисовываться типичные клетки. Только после рождения начинается и быстро происходит развитие нормальной слизистой, благодаря интенсивной резорбции миксоидной ткани.
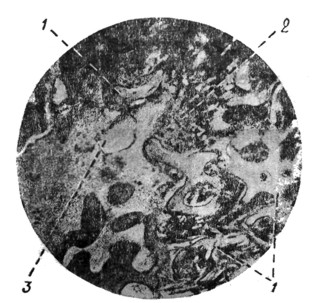 |
Рис. 28. Микрофотография уха ребенка восьми месяцев (по Комендантову). |
К моменту рождения миксоидная ткань заполняет три главные полости среднего уха: собственно барабанную полость, antrum и recessus. Иногда - в виде тяжей и сплетений - ткань эту находят в полостях среднего и внутреннего уха детей и даже стариков (рис. 28). Резорбция миксоидной ткани вызывается многими причинами: воспалительными явлениями, дыхательными движениями новорожденного, биологическими силами организма, пульсаторными движениями сосудов, прилегающих к среднему уху, - в частности луковицы яремной вены. Воспалительные процессы, по мнению некоторых исследователей, влияют на развитие разных типов строения слизистой оболочки. Различают: фиброзную, гиперпластическую и смешанную фиброзно-гиперпластическую слизистую. Результатом такой вариации слизистой оболочки некоторые авторы считают и различный процесс пневматизации сосцевидного отростка, образование пневматических клеток которого начинается на втором году жизни ребенка, достигая верхушки отростка к концу 3 или 4 года жизни. Различное распространение сети ячеек, конфигурации их, четкость их разграничения, различное строение слизистой, их выстилающей, степень развития сосудистой системы и иннервации составляют сущность вариантов конституции сосцевидного отростка (рис. 29).2
Сосцевидный отросток развивается из pars mastoidea новорожденного ребенка только к концу первого года его жизни. К этому моменту развивается уже m. sterno-cleido - mastoideus как важный фактор в функции поворота головы и дыхания, содействующий своею работою росту сосцевидного отростка.
Особенности уха раннего детского возраста состоят в следующем.
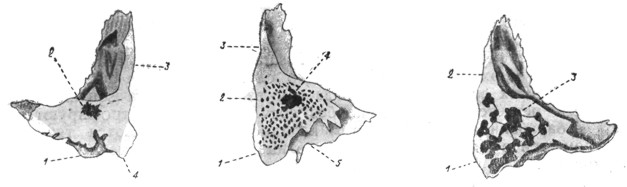 |
||
Рис. 29. Варианты сосцевидных отростков. | ||
|
|
|
Костной части наружного слухового прохода у них нет, есть только annulus tympanicus.
Сосцевидного отростка у детей нет, есть только краешек pars petrosa, к которому прикреплен га. sterno-cleidomastoideus, заставляющий его развиваться в длину и ширину благодаря работе, осуществляемой в связи с вертикальным положением, принимаемым ребенком в течение своего роста.
Евстахиева труба у детей широко открыта, дает доступ воздуху в барабанную полость и в antrum, где вызывает резорбцию слизистой, оказывая существенное влияние на пневматизацию медленно развивающегося сосцевидного отростка.
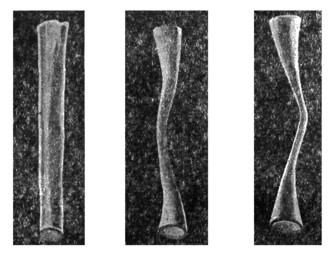 |
Рис. 30. Варианты Евстахиевых труб. |
Fossa subarcuata у детей еще полна миксоидной ткани, исчезающей лишь впоследствии.
Имеются незаросшие еще швы; fissura squamo-mastoidea открыта; через нее проходят сосуды, нередко служащие путем распространения ушных осложнений. Иногда и у взрослых fissura squamo-mastoidea остается открытой, но без сосудов, которые атрофируются. В барабанной полости от миксоидной ткани остаются тяжи и перемычки в виде складок, нередко вызывающих врожденную тугоухость.
И генотипические и фенотипические факторы влияют на развитие различных анатомических вариантов наружного, среднего и внутреннего уха; отчасти отсюда вытекают различные физиологические варианты и степени слуховой и статической одаренности индивидуумов, с одной стороны, а с другой - различная реакция составных частей уха на патологические факторы, т.е. разное течение болезни не только у различных индивидуумов, но даже у одного и того же человека в разные периоды его жизни и в различной трудовой обстановке.
Изложенное составляет элемент конституциональной основы возрастной и профессиональной патологии в отиатрии.
|
The requested URL /down.htm was not found on this server.