
The requested URL /topic.htm was not found on this server.
Общим морфологическим свойством органа обоняния у всех животных, как позвоночных, так и беспозвоночных, является наличие первичных чувствующих (обонятельных) клеток. От базилярной части такой клетки отходит отросток, непосредственно переходящий в нервное волокно, направляющееся к ганглию или к мозгу, а периферическая, свободная часть клетки снабжена специфическими волосками, воспринимающими раздражение. Между чувствующими клетками расположены индифферентные опорные клетки, которые, впрочем, у ряда беспозвоночных отсутствуют. В наиболее примитивном виде орган обоняния представляет собой выпячивание эктодермы на поверхности тела в форме так называемой обонятельной ямки, выстланной простым или мерцательным эпителием, содержащим и обонятельные клетки.
Органы обоняния беспозвоночных животных чрезвычайно разнообразны по своему строению и локализации, у различных типов и классов не являются гомологичными образованьями и совершенно не схожи с органами обоняния позвоночных. Вообще, нет достаточно убедительных доказательств того, что так называемые обонятельные органы беспозвоночных действительно выполняют во всех случаях обонятельную функцию. Уже у кишечнополостных (медузы), у многих червей и моллюсков на различных участках поверхности тела имеются обонятельные ямки, выстланные мерцательным эпителием с обонятельными клетками. У моллюсков, кроме того, появляются обонятельные щупальцы и осфрадии (шпенгелевы органы), соединяющиеся, как и обонятельные ямки у червей, с головным нервным узелком. Осфрадии у водных моллюсков расположены в полости мантии у основания жабер и в простейшем виде представляют утолщенный эпителий, а в более сложной форме имеют гребенчатый вид, напоминающий жабры. У легочных моллюсков осфрадии в виде двух ямок помещаются вблизи дыхательного отверстия. Обонятельные щупальцы у наземных моллюсков находятся на голове.
Совершенно особенное и разнообразное строение имеют органы обоняния членистоногих; хитиновый панцирь не оставляет места на поверхности тела для этих нежных образований; здесь обонятельные волоски - окончания чувствующих клеток - расположены на сяжках. У раков органом обоняния считают коническое возвышение на основном членике наружной пары сяжков. Свободный конец этого возвышения или покрыт тонкой оболочкой, или снабжен маленьким отверстием, которое ведет внутрь, где открывается железистый орган ("зеленая железа"), лежащий в головогруди. У насекомых, вероятно, сяжки являются также органами для распознавания состояния окружающей среды: влажности воздуха, его температуры и т. д.
Орган Обоняния у всех хордовых животных является гомологичным, хотя функция его у различных классов и не оказывается совершенно тождественной. У наземных животных орган обоняния воспринимает раздражения от газообразных веществ или от веществ, взвешенных в воздухе, а у рыб - от растворенных в воде веществ, т. е. приближается к органу вкуса. Впрочем, существует мнение, что и у рыб имеется какая - то качественная разница между ощущениями вкуса и обоняния. С другой стороны, даже у человека в некоторых отношениях нельзя провести четкую грань между вкусом и обонянием.
Несколько изолированное положение среди хордовых занимает обонятельный орган ланцетника, имеющий форму одиночной плоской ямки с мерцательным эпителием, расположенной на левой стороне переднего конца тела животного и прямо соединенной с передним концом центральной нервной системы.
У черепных (позвоночных) животных орган обоняния развивается в виде парного утолщения эктодермы, на переднем конце головы (обонятельные плакоды), которое затем погружается глубже, образуя сначала простую обонятельную ямку. Исключением оказываются только круглоротые рыбы, имеющие непарный орган обоняния. В простейшем виде обонятельный орган представляется в виде парных ямкообразных углублений кожи, выстланных чувствующим эпителием и помещающихся на переднем конце головы. Обонятельный эпителий состоит из длинных тонких первично чувствующих клеток и не менее длинных опорных клеток. У обонятельных клеток образуется веретенообразное вздутие на месте ядра, на периферическом конце имеются пучки чувствующих волосков, а со стороны проксимального конца исходят волокна обонятельного нерва. У рыб в обонятельном органе, кроме того, встречаются почки, похожие на вкусовые.
Орган обоняния позвоночных, как уже упомянуто, представляет два отличных типа: непарный орган у круглоротых рыб (Monorhini) и парный орган у прочих позвоночных (Amphirhini). По мнению Геккеля и Купфера, непарная обонятельная ямка круглоротых может быть сравнена с непарной же ямкой ланцетника, т. е. является одиночной по своему первоначальному происхождению. На основании других взглядов (Gegenbauer, Dubosch, Peter), подобная непарность органа обоняния у круглоротых представляет собой вторичное явление, возникшее в результате слияния парных органов; в пользу такого взгляда говорит наличие у круглоротых рыб парного обонятельного нерва.
Парные обонятельные ямки - плакоды-в их первоначальной форме открываются каждая одним отверстием наружу. У многих же рыб первоначально (у зародыша) имеется одиночное отверстие, которое потом делится на два - переднее и заднее, и в результате каждая обонятельная ямка получает по два наружных отверстия. Таким образом, создается первоначальное приспособление к обмену воды в обонятельной ямке: она теперь уже имеет форму канала с двумя отверстиями (ноздри), способствующими сквозному протеканию через нее воды и, следовательно, усилению обонятельной способности. У селяхий обонятельные ямки получают новое приспособление для обмена воды - желобок, соединяющий ямку, которая расположена непосредственно впереди рта, с ротовой полостью; желобок, в свою очередь, прикрыт снаружи кожной складкой и превращается как бы в трубочку. Dipnoi, Amphibia и Amniota имеют вместо желобка сформировавшийся канал (за счет срастания краев кожной складки, прикрывающей желобок у селяхий), соединяющий обонятельные ямки с полостью рта. Отверстия, соединяющие обонятельный орган с ротовой полостью, носят название хоан, задних носовых отверстий. У Amniota эмбриологический процесс развития носовой полости повторяет на известном этапе жизни зародыша стадию желобка, имеющегося у селяхий.
Обонятельные ямки у позвоночных защищаются снаружи скелетной капсулой, срастающейся с передней частью мозговой коробки.
У наземных позвоночных орган обоняния, возникший еще в условиях водной жизни, оказывается снабженным большим количеством слизеотделительных желез, обеспечивающих влажное состояние обонятельного эпителия. Кроме того, в носовой полости наземных животных (начиная с амфибий) обособляются два участка: один из них - pars respiratoria - служит только для проведения воздуха, и его эпителий не содержит обонятельных клеток; другой - pars olfactoria - является обонятельной областью.
Обонятельная способность оказывается хорошо выраженной у некоторых рыб, менее - у амфибий, рептилий и птиц и снова более мощно развитой у млекопитающих. Можно проследить параллелизм между развитием обонятельной способности и существованием кожных желез, выделяющих пахучие вещества.
У всех позвоночных в органе обоняния отмечается наличие структурных приспособлений, увеличивающих поверхность обонятельной полости. У водных животных это происходит путем образования многочисленных мелких радиальных складок, а у наземных - путем образования в обонятельной полости более крупных складок, снабженных скелетным остовом (так называемые раковины). Наибольшей сложности система раковин достигает у млекопитающих. Увеличение поверхности у наземных животных наблюдается не только в обонятельной части носовой полости, но также и в дыхательной области. Оно осуществляется за счет развития складок и раковин в просвете носовой полости, а также путем разрастания последней в соседние кости в виде придаточных пазух. Последние, резко увеличивая общую поверхность всей системы носовой полости, как правило, не содержат в своей слизистой оболочке обонятельных клеток.
Далее, у наземных строение носовой полости усложняется развитием слезноносовых каналов и обособлением парного участка носовой полости в так называемый якобсонов орган, открывающийся потом в ротовую полость.
Круглоротые рыбы имеют непарный обонятельный орган, который вторично соединяется с гипофизарной трубкой и через нее открывается наружу в виде непарной "ноздри" на спинной стороне головы. При сжиманиях и растяжениях гипофизарного мешка по гипофизарному каналу через "ноздри" в орган обоняния проходит ток воды. У миксин гипофизарная трубка другим своим концом открывается также и в ротовую полость; в этом случае она проводит воду не только к обонятельному органу, но и к жабрам; таким образом, достигается постоянное обновление воды в органе обоняния.
Обонятельный орган круглоротых имеет форму мешка. На его внутренней поверхности выявляются радиальные (меридиональные) складки. На складках и в углублениях между ними располагается обонятельный эпителий, переходящий в пару обонятельных нервов. Наличие складок значительно увеличивает обонятельную поверхность. У миног мешок разделяется на две части вертикальной складкой. Обонятельный мешок круглоротых окружен снаружи хрящевым выступом черепа или хрящевой носовой капсулой.
Связь обонятельного органа с гипофизом и непарность обонятельного органа характерны для круглоротых; у других позвоночных обонятельный орган не имеет никакого отношения к гипофизу, является парным и открывается наружу двумя ноздрями.
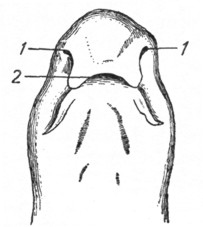 |
Рис. 1. Нижняя поверхность головы акулы (по Видерсгейму). |
Селяхии. Парные обонятельные ямки селяхий (поперечноротых) лежат на нижней поверхности выдающегося вперед рыла, впереди рта, иногда на значительном расстоянии от него, и сообщаются желобками с ротовой полостью (рис. 1, 2). У скатов и акул желобок прикрывается сверху кожной клапанообразной складкой и как бы превращается в трубочку, открывающуюся под верхнюю губу. Вода, попадая через ноздри в носовые полости, выходит из них через указанные желобки в полость рта. Внутренняя поверхность носовой полости еще более увеличивается за счет развития на первоначальных обонятельных складках более мелких, вторичных и даже третичных складок.
Осетровые и костистые рыбы. У ганоидов и костистых рыб обонятельный орган состоит из двух мешков, из которых каждый открывается наружу двумя отверстиями, расположенными на верхней стенке черепа (рис. 3). Обонятельный орган, таким образом, не находится в связи с ротовой полостью, как это имеет место у селяхий и наземных позвоночных. В процессе развития обонятельный орган, первоначально располагающийся на вентральной поверхности головы, перемещается в латеральном направлении на ее дорсальную поверхность. Из двух отверстий, которыми каждый обонятельный мешок открывается наружу, переднее обычно помещается на щупальцевидном отростке. Внутри на стенках обонятельного органа расположены увеличивающие его поверхность радиальные складки. Нередко обонятельная полость имеет очень сложное строение, так как основной обонятельный мешок стоит в связи с рядом дополнительных обонятельных мешков. Например, особенно сложными оказываются обонятельные органы у ганоидов Polypterus. Они глубоко заходят в хрящевой череп и состоят из преддверия и собственно обонятельной ямки, которая, в свою очередь, оказывается разделенной радиальными перегородками на 6 полостей; в каждой из них расположены многочисленные веерообразно расходящиеся складки, заключающие в себе ветви обонятельного нерва. У подотряда сростночелюстных рыб, например у рода Tetrodon, отмечается редукция органа обоняния. Здесь нет обонятельных отверстий, но вместо них имеются плотные щупальцы, в толще которых помещается обонятельный нерв и на поверхности которых расположены скопления обонятельных клеток. У некоторых рыб не оказывается и щупальцев; в таких случаях обонятельный нерв оканчивается в области небольшого пигментированного участка кожи, а обонятельная, полость вовсе отсутствует.
Двоякодышащие рыбы. Двоякодышащие составляют небольшую группу вымирающих животных, которые соединяют в себе черты организации рыб и амфибий. Орган обоняния у них (рис. 4) представляет как бы дальнейшее развитие и усложнение органа обоняния селяхий. Носовая полость имеет не только ноздри, но уже и задние отверстия - хоаны, открывающиеся в ротовую полость. Наружные отверстия (ноздри) здесь также отличаются особенностью в расположении: у зародыша они лежат снаружи (как и у селяхий), но затем вследствие срастания верхней губы с верхней челюстью перемещаются внутрь ротовой полости, открываясь под верхней губой. Таким образом, обонятельный орган открывается в рот обоими отверстиями (передним и задним). Внутри обонятельного органа имеются продольные складки, соединенные между собой поперечными. Обонятельный орган, по-видимому, обладает способностью воспринимать ощущения не только из воды, протекающей через его полость, но и из воздуха. Двоякодышащие рыбы, помимо водного дыхания жабрами, заглатывают также и воздух для наполнения легких. При указанной связи обонятельного органа с ротовой полостью заглатываемый ртом воздух проходит и через орган обоняния. Как известно, двоякодышащие населяют водоемы, которые в жаркое время года сильно высыхают; поэтому, например, австралийский чешуйчатник (Ceratodus Forsteri), спасаясь летом в ямах, остающихся при высыхании водоемов, пользуется легочным дыханием, так как вода уже оказывается непригодной для дыхания. Южноамериканские и африканские (Protopterus) виды двоякодышащих при наступлении сухого времени года зарываются в ил и образуют вокруг своего тела кокон, выстланный изнутри слизью и имеющий открытый наружу канал, по которому от наружной поверхности до рта проходит необходимый для дыхания воздух. В таком состоянии имеет место исключительно легочное дыхание через рот, и при расположении обоих носовых отверстий в ротовой полости воздух протекает также через орган обоняния. Слизистые железы омачивают поверхность обонятельных мешков и предохраняют их от высыхания. У двоякодышащих впервые возникает связь обоняния с воздушным дыхательным аппаратом.
 |
 |
Рис. 2. Поперечный разрез через обонятельную область головы акулы (по Шмальгаузену). |
Рис. 3. Голова осетровой рыбы (по Видерсгейму). Видны переднее и заднее носовые отверстия. |
Амфибии. Обонятельный орган амфибий представляет новые особенности. Обонятельный мешок имеет ноздрю, открывающуюся на спинной части головы, и первичную хоану, открывающуюся на крыше ротовой полости, причем полость обонятельного органа оказывается приспособленной и для новой функции - прохождения воздуха при дыхании легкими. Таким образом, приспособление к обмену воздуха в органе обоняния привело к усовершенствованию органов дыхания; орган обоняния стал участвовать в акте дыхания, и это отразилось на его дальнейшей эволюции. Первичную хоану амфибий нельзя просто сравнивать с задним носовым отверстием рыб: она возникает не в результате врастания в глубину носового желобка, но путем прорыва обонятельного мешка в энтодермальную переднюю кишку. Первичные хоаны лежат у постоянножаберных еще внутри губ, а у прочих амфибий отступают назад; у лягушек и саламандровых они отграничены верхней челюстью и небной костью.
Кроме наличия внутренних носовых отверстий, характерной особенностью обонятельных органов амфибий является образование слезноносового канала, отсутствующего только у некоторых низших форм (постоянно живущих в воде), а также возникновение у наземных животных специальных желез, открывающихся в носовую полость. У низших форм амфибий еще сохраняются общие с двоякодышащими черты строения обонятельного органа: носовая полость имеет вид простого канала, покрытого на медиальной стороне обонятельным эпителием, а на латеральной стороне - индифферентными клетками. Канал открывается в ротовую полость хоаной, которая иногда с медиальной стороны прикрыта маленькой складкой. Внутренние стенки органа снабжены узкими радиальными складками, которые у выше стоящих форм уже отсутствуют.
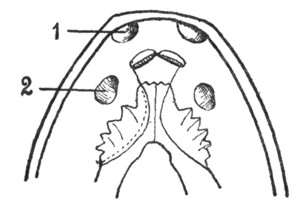 |
Рис. 4. Небо двоякодышащих - Ceratodus (по Холодковскому). |
У более высоко организованных амфибий отмечаются некоторые особенности, приближающиеся к строению амниотных. Длинная складка неба прикрывает с латеральной стороны задние носовые отверстия и отграничивает их в виде первичных хоан от ротового свода, как бы погружая их в глубину. Эта складка оказывается первым намеком на вторичное небо, которое приобретает большое значение у других четвероногих. На внутренней выстилке обонятельной полости также происходят изменения: вместо узких складок на стенках появляются широкие валики (прототип носовых раковин), отделяющие от основной полости обособленные слепые отделы. Так создается система воздушных полостей, в которой у большинства амфибий можно различать два отдела - верхний и нижний (рис. 5). В зависимости от воздушного дыхания в носовой полости обособляются: pars respiratoria - наружные отделы органа, не содержащие обонятельных клеток, и pars olfactoria - медиальные отделы, покрытые обонятельным эпителием.
Заслуживает внимания отмечающийся у ряда амфибий на медиально - вентральной стенке носовой полости ответвляющийся нижний слепой мешок - зачаток якобсонова органа амниотных. Он обособляется уже у некоторых хвостатых амфибий в виде желобка, расположенного на внутренней поверхности нижнего отдела носовой полости и выстланного обонятельным эпителием. У хвостатых постоянножаберных первый прототип якобсонова органа имеет вид небольшой полоски чувствительного эпителия, расположенной в тех же участках. У саламандровых и бесхвостых он выражен более отчетливо и наиболее высокого развития достигает у безногих амфибий, у которых он вполне обособляется и имеет вид слепого мешка, лежащего под носовою полостью и сообщающегося с нею в задних отделах. Одновременно с развитием якобсонова органа обособляется и соответствующая часть обонятельного нерва. Слизистые железы, открывающиеся в носовую полость наземных амфибий, расположены снаружи от обонятельной полости (верхняя носовая железа), кнутри от нее (нижняя или якобсонова носовая железа) и у заднего края хоан (небная железа).
Переходя к рассмотрению органа обоняния у Amniota (рептилии, птицы, млекопитающие), следует отметить у них большую потребность в воздухе. Носовая полость приспособляется к его очистке, увлажнению и согреванию. Обе носовые полости оказываются сближенными. Если у рассмотренных выше классов позвоночных они отделены друг от друга обширными межуточными тканями, то у Amniota они разделяются лишь тонкой носовой перегородкой.
Строение обонятельного органа у группы Sauropsida (рептилии и птицы) имеет известную общность у отдельных входящих в эту группу классов; у млекопитающих же отмечаются значительные особенности, усложняющие строение обонятельного органа.
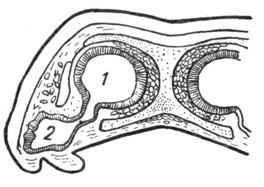 |
Рис. 5. Поперечный разрез носовой полости лягушки (по Гегенбауеру). |
Рептилии. Среди основных групп рептилий (ящерицы, змеи, черепахи, крокодилы) в строении обонятельного органа имеются некоторые варианты, что зависит от образа жизни, а также от формы головы животного. Например, высокая голова черепахи имеет короткую, но высокую носовую полость, а длинная и низкая голова крокодила - вытянутую в длину, но уплощенную носовую полость. Наружные носовые отверстия у крокодилов и у живущих в воде змей закрываются особыми клапанами, которых нет у других рептилий. В то время как ноздри почти всегда лежат вблизи конца рыла, задние носовые отверстия то направлены к небу прямо вниз от дна носовой полости (змеи, ящерицы), то в виде канала идут далеко назад (крокодилы). В качестве примера типичного строения носовой полости у рептилий можно рассмотреть носовую полость ящерицы (рис. 6). Носовая полость разделяется на две функционально различные области: выше расположена pars olfactoria и ниже - pars respiratoria. Последняя дифференцирована на три отдела: начинающийся от ноздрей передний отдел или преддверие (vestibulum), выстланное плоским эпителием, средний отдел (или основная полость), наиболее просторный по объему и соединяющийся с обонятельной областью, и задний отдел (ductus nasopharyngeus), открывающийся хоанами в полость рта. На латеральной стенке среднего отдела у рептилий имеется челюстная раковина (concha seu maxilloturbinale), состоящая из покрытого слизистой оболочкой скелетного остова (хрящ или кость) в виде слегка завитого выступа; она подразделяет носовую полость на две вышеупомянутые области - обонятельную и дыхательную. Таким образом, просвет среднего отдела оказывается в значительной степени суженным, а его поверхность - увеличенной. Раковина, отчетливо выраженная у ящериц и змей, оказывается в зачаточном состоянии в виде плоского валика у черепах и особенно сильно развитой бывает у крокодилов, у которых позади основной раковины имеется еще псевдораковина. У крокодилов, кроме того, выявляется уже придаточная пазуха носа (sinus paranasalis), сообщающаяся с основной носовой полостью, выстланная также слизистой оболочкой, но не содержащая обонятельных клеток. Слезносовой канал открывается под раковину, в нижнюю часть носовой полости. Задние носовые отверстия, как уже упомянуто, могут занимать различное положение. У ящериц и змей они, как и у амфибий, представляют собой "первичные хоаны", открывающиеся на крыше ротовой полости и лишь отчасти прикрытые боковыми небными складками. У черепах и крокодилов эти складки оказываются взаимно сросшимися, превратившимися во вторичное твердое небо, и носовые полости уже не открываются в рот первичными хоанами, а имеют "вторичные хоаны", сильно отодвинутые кзади (особенно у крокодилов) и открывающиеся у заднего конца твердого неба. Подобное отодвигание хоан назад, ближе к глотке и гортани, дает возможность животному свободно дышать носом даже в то время, когда во рту находится пища.
Эпителий носовой полости у рептилий приобретает характер слизистой оболочки, поддерживаемой во влажном состоянии секретом бокаловидных клеток и сложных желез. Различаются железы верхние (или наружные), лежащие внутри раковины и открывающиеся на границе преддверия и носовой полости, и нижние (или внутренние), открывающиеся около носовой перегородки.
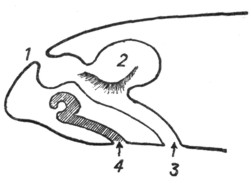 |
Рис. 6. Схема сагиттального разреза через обонятельные органы ящерицы (по Видерсгейму). |
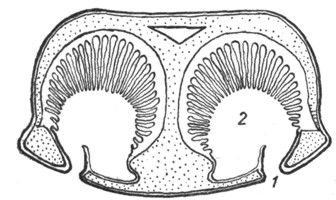
Якобсонов орган у черепах и крокодилов имеется лишь в эмбриональном периоде в виде желобка у нижнего края медиальной стенки носовой полости. Напротив, у ящериц и змей он сохраняется в течение всей жизни. У них якобсонов орган (рис. 7) нацело оказывается отделенным от полости носа и получившим сообщение с ротовой полостью. Этот орган имеет форму трубки, оканчивающейся слепым концом под нижнемедиальным отделом носовой полости, и посредством канала сообщается с полостью рта. Эпителиальная выстилка якобсонова органа снабжена обонятельным эпителием, а стенка содержит Под носовыми полостями (1) расположены якобсоновы органы, устья которых (2) открываются в полость рта. хрящевую капсулу. Орган снабжен ветвями обонятельного и тройничного нервов. В просвет органа вдается небольшой имеющий хрящевую основу выступ, который может быть уподоблен раковине. Функция якобсонова органа точно не известна; по-видимому, он служит для восприятия обонятельных ощущений от находящейся во рту пищи. В отличие от наполненной воздухом носовой полости якобсонов орган оказывается постоянно наполненным жидким секретом якобсоновых желез. Это обстоятельство дает основание предполагать функциональную разницу между обонятельной областью носовой полости и якоб-соновым органом. Среди ящериц якобсонов орган отсутствует у хамелеонов, а среди змей оказывается особенно сильно развитым у ведущих подземный образ жизни слепунов (Typhlopidae), у которых носовая полость редуцирована.
Птицы. Орган обоняния у птиц по сравнению с рептилиями имеет более сложное внутреннее устройство (рис. 8, 9). Наоборот, строение неба более примитивно. Наружные носовые отверстия обычно помещаются на корне клюва; оба они иногда могут слиться в одно (у буревестников); иногда на некотором протяжении у входа в носовую полость недостает носовой перегородки, отчего образуются nares perviae. Носовая полость на своей латеральной стенке имеет уже не одну, как у рептилий, а три раковины. Прежде всего в преддверии расположена раковина (concha vestibuli), не имеющая гомолога у других амниотных; она покрыта плоским эпителием и не имеет обонятельного значения; очевидно, ее функция заключается в обогревании и увлажнении вдыхаемого воздуха, а также в защите носовой полости от проникновения инородных тел. Средняя раковина, расположенная в дыхательной области носовой полости, гомологична раковине рептилий; у птиц она более мощно развита и изогнута в виде спирали. Эта челюстная раковина (maxilloturbinale) также не имеет обонятельной функции, но представляет хороший фильтр для вдыхаемого воздуха. Третья верхняя раковина (nasoturbinale) расположена выше и несколько кзади от челюстной; она является выступом наружной стенки обонятельной области и сама покрыта обонятельным эпителием. Выступ латеральной стенки, оформляющий данную раковину, содержит воздухоносную бухту, имеющую сообщение с челюстной придаточной пазухой. Верхние раковины более сильно развиты у хищных птиц, средние раковины - у куриных и передние - у певчих. У птиц хорошо развита система воздухоносных придаточных пазух носа. Якобсонов орган отсутствует; его зачатки можно найти только у эмбриона.
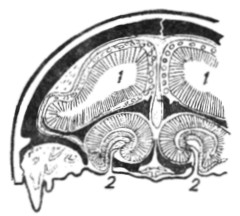 |
Рис. 7. Поперечный разрез через голову медяницы (по Гегенбауеру). |
В носовые полости открываются носовые железы, расположение которых бывает очень различным; наиболее часто они помещаются на лобной кости. Вторичные небные складки у птиц не срастаются (хотя иногда почти соприкасаются) и не приводят к образованию вторичного неба; поэтому на своде рта носовая полость открывается щелевидными первичными хоанами. В общем, у птиц обоняние не играет большой роли и основным из органов чувств является зрение.
Млекопитающие. Обоняние у млекопитающих имеет гораздо большее значение, чем у амфибий, рептилий или птиц. Только вторичным путем, главным образом в связи с развитием глаз и мозга, у млекопитающих может происходить уменьшение роли обоняния. Однако, и в последних случаях носовая полость как часть органа дыхания, обеспечивающая проведение воздуха, оказывается значительно более сложно построенной по сравнению с носовой полостью других позвоночных. У зверей обоняние играет большую роль для ориентации в пространстве, в отыскивании пищи, в оберегании от врагов, в половой жизни и т. д. Наиболее слабо развита обонятельная способность у приматов и водных млекопитающих (ластоногие и китообразные); среди последних обонятельный орган наиболее редуцирован у зубастых китов. Эта особенность стоит в связи с водным образом жизни. Все млекопитающие по степени развития обонятельного органа могут быть подразделены на три группы: аносматические формы (зубастые киты), микросматические формы (беззубые киты, ластоногие и приматы) и макросматические формы (все остальные млекопитающие). Большое развитие решетчатой кости у млекопитающих создает новое расположение органа обоняния: в то время как у птиц он лежит перед глазницами, у млекопитающих носовая полость представляется более объемистой, простирается кзади между обеими глазницами и отграничивается пластинкой решетчатой кости сверху и кзади от дна черепномозговой полости. Наружное носовое отверстие ведет в преддверие носа (выстланное плоским эпителием), которое без резкой границы переходит в носовую полость. В носовой полости имеется система раковин. Из них maxilloturbinale делит всю носовую полость на два отдела: верхний - обонятельный и нижний - дыхательный, причем увеличение обонятельной поверхности и верхнего отдела происходит путем развития зтмоидальных раковин. Дыхательная часть получает иннервацию только от тройничного нерва. Обе носовые полости отделяются одна от другой носовой перегородкой, состоящей из перпендикулярной пластинки решетчатой кости, сошника и в передних отделах - из хряща. Хрящи могут входить также в состав скелета наружного носа (как остаток первоначальной хрящевой обонятельной капсулы) и даже иногда создавать трубкообразную форму его (например у землероек, медведеобразных хищников). У ныряющих животных отмечается наличие заслончатого аппарата для закрытия ноздрей, а иногда (у морских собак) он заменяется особым запирательным мускулом. У других животных удлинение хрящевых подпорок наружного носа приводит к тому, что он приобретает хоботообразную форму, например у крота, тапира, свиньи и всего более у слона, где хобот является одновременно органом осязания и хватания. Мускулы, приводящие в движение крылья носа оказываются более развитыми при существовании заслонок и особенно мощно выражены при хоботообразных формах носа. Они либо начинаются от верхней челюсти и тянутся вдоль всего хобота, либо расположены в виде коротких поперечных и продольных мышечных пучков, число которых в хоботе слона особенно велико. У китообразных к наружным отверстиям носа примыкают брызгательные каналы, сообщающиеся с брызгательными мешками; через эти отверстия киты выпускают свой "фонтан". Ноздри у китообразных отодвинуты от конца рыла далеко кзади и открываются на лобной области головы; такое положение ноздрей дает возможность животному дышать, когда рыло находится уже под водою. У китов имеются две ноздри, а у дельфинов - только одна. У роющих животных (крот, свинья) на дистальной части носа развивается окостенение (os praenasale), существующее, впрочем, и у некоторых других млекопитающих (хоботных).
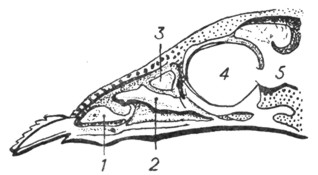 |
Рис. 8. Сагиттальный распил головы птицы (по Бютчли). |
Носовые раковины состоят из упомянутой максиллярной раковины (maxilloturbinale seu concha inferior в анатомии человека), затем свойственной многим млекопитающим nasoturbinale (гомолог которой имеется у птиц), расположенной над нижней раковиной, и из системы раковин решетчатой кости (ethmoturbinalia). Maxilloturbinale и nasoturbinale представляют собой выросты на наружной стенке носовой полости. Обе эти раковины простираются вдоль всей передней части носовой полости и своими передними концами иногда доходят до наружного отверстия, тогда как решетчатые раковины лежат более глубоко в задней части носовой полости. Максиллярная раковина может иметь различную форму: то она является простым завитком, то на свободном своем крае образует двойной завиток, закручивающийся одной лопастью кверху, а другой-книзу; у приматов она имеет более простое строение. Эта раковина оказывается исключительно дыхательной, так как лишена обонятельного эпителия. Nasoturbinale также в основном является дыхательной раковиной, и обонятельный эпителий имеется лишь в самом заднем ее отделе. У обезьян и человека nasoturbinale редуцирована до степени небольшого выступа на латеральной стенке носовой полости (так называемый носовой вал - agger nasi).
 |
Рис. 9. Поперечный разрез носовой полости птицы (по Видерсгейму). |
Этмоидальные раковины расположены на внутренней поверхности латеральной пластинки решетчатой кости и направляются к ее срединной вертикальной (перпендикулярной) пластинке, входящей в состав носовой перегородки. Покрывающая этмоидальные раковины слизистая оболочка содержит обонятельные клетки. Число этмоидальных раковин может быть различным: чаще всего их бывает 3; у сумчатых, насекомоядных - до 5; у копытных количество раковин доходит до 8, а у приматов уменьшается до 2; впрочем, у зародыша человека их бывает гораздо больше. Описанные раковины относятся к группе так называемых endoturbinalia, т. е. их завитки вследствие большого размера базальной части раковин лежат ближе к срединной линии. В промежутках между endoturbinalia располагаются более мелкие по размеру раковины, у которых базальная пластинка имеет меньшее протяжение. Эти раковины не видны при осмотре боковой стенки носовой полости на сагиттальном распиле (рис. 10), но их можно обнаружить на фронтальном распиле (рис. 11). Они называются прикрытыми раковинами (conchae obtectae) или, поскольку они располагаются более кнаружи, - ectoturbinalia. Количество прикрытых раковин также варьирует, причем у человека ectoturbinalia отсутствуют и их рудиментом является bulla ethmoidalis. Перемена положения головы и развитие лобных долей мозга у приматов приводят к перемещению положения в пространстве и носовых раковин: этмоидальные раковины располагаются в носовой полости не кзади, а над maxilloturbinale. Таким образом, у человека нижняя раковина соответствует maxilloturbinale, а средняя и верхняя - двум этмоидальным раковинам. У макросматических млекопитающих обонятельный эпителий покрывает почти всю поверхность обонятельных раковин, у человека же область его распространения ограничивается только задне-верхним отделом носовой полости. Еще более рудиментарной оказывается обонятельная область у китообразных и полностью отсутствует у зубастых китов. У других не постоянно живущих в воде животных (утконос, бобер, тюлень) обонятельная область имеет очень простое строение, раковины развиты слабо. Однако, иногда у довольно близко стоящих форм, например у ехидны, у утконоса, наблюдаются различные степени развития этмоидальных раковин: у утконоса они редуцированы, но сильно развиты у ехидны.
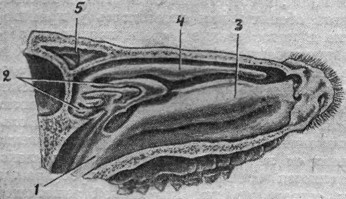 |
Рис. 10. Сагитальный разрез носовой полости теленка (по Peter). |
Носовая слизистая оболочка богата мелкими железами. Кроме того, у многих млекопитающих имеются крупные боковые железы, гомологичные боковым железам рептилий; реже бывают подобные железы и в носовой перегородке. Особенно много больших желез у грызунов; эти железы отчасти выходят кнаружи из носовой капсулы и орошают носовую полость, небноносовой канал и наружный нос.
Уже у крокодилов и птиц носовая полость оказывается увеличенной в объеме благодаря образованию воздухоносных полостей в соседних костях. Особенно сильно развитой бывает система придаточных пазух у млекопитающих. Наиболее постоянно придаточные пазухи располагаются в верхнечелюстной кости (sinus maxillaris, seu antrum Highmori - гайморова пещера человека), в лобной (sinus frontalis) и основной (sinus sphenoidalis) костях, причем иногда в лобной и основной пазухах могут быть и обонятельные валики, но в подавляющем большинстве случаев придаточные пазухи не выполняют обонятельной функции и являются исключительно воздухоносными полостями. Иногда пазухи располагаются и в других костях, заходя (например у слона) в теменные и височные кости. Основное значение придаточных пазух, невидимому, заключается в облегчении веса черепа, так как они особенно сильно развиты у животных, имеющих массивный, отягощенный рогами или жевательным аппаратом череп, который поддерживается впереди туловища силой затылочных мышц и связок, и наоборот, слабо развиты у мелких зверей (грызуны) и у животных, живущих в воде (тюлени, киты). Расположение, число и величина придаточных пазух, таким образом, подвержены у различных отрядов и семейств млекопитающих большим колебаниям (см. также главу "Физиология").
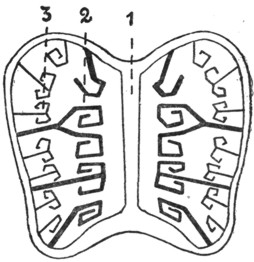 |
Рис. 11. Схема поперечного разреза носовой полости млекопитающего (козы) (по Шимкевичу). |
Придаточные пазухи обычно появляются и развиваются не столько в период эмбрионального развития животного, сколько уже при внеутробной жизни, и пневматизируются как бы под влиянием врастания слизистой оболочки носовой полости в окружающую ее костную ткань. Поэтому у разных отрядов получаются различия в размерах и распространении придаточных пазух: одна и та же пазуха (например лобная) у одних животных (хищные) помещается только в лобной кости, а у других (жвачные) врастает в теменные и даже затылочные кости. И наоборот, у различных животных одна и та же кость оказывается в разной мере пневматизированной в зависимости от степени развития придаточных пазух. Основная пазуха развивается из заднего конца носовой полости, лобная - из верхнего отдела и челюстная пазуха - из промежутка между челюстной и первой решетчатой раковинами. У копытных, многих плотоядных и приматов выражена, кроме того, система мелких воздухоносных полостей в решетчатой кости (cellulae ethmoidales), которые возникают из щелей между обонятельными раковинами и заходят также в другие соседние кости. У приматов придаточные пазухи развиты гораздо слабее, чем, например, у копытных или жвачных; у некоторых обезьян (гиббон) отсутствуют лобные пазухи или даже все пазухи за исключением челюстной (игрунковые обезьяны).
Якобсонов орган у млекопитающих представлен различно. У яйцеродных сумчатых, неполнозубых, копытных, насекомоядных и грызунов имеется с каждой стороны в носовой перегородке по якобсонову органу в виде длинной сагитально лежащей трубочки, окруженной хрящевой капсулой (рис. 12); здесь орган оканчивается слепо, а спереди открывается либо в ротовую полость, проходя через canalis nasopalatinus, либо открывается в самой же носовой полости (копытные, грызуны). У яйцеродных сохраняется свойственная Sauropsida раковина на наружной стенке органа. Особенно сильно развит якобсонов орган у некоторых летучих мышей и у яйцеродных млекопитающих. Эпителиальная выстилка органа содержит обонятельный эпителий. Иннервируется орган обонятельным и тройничным нервами. Просвет органа никогда не содержит воздуха, но заполнен жидкостью. У человека якобсонов орган обычно редуцируется на 9-м месяце утробной жизни и только в виде редкого исключения может сохраниться canalis nasopalatinus и даже весь орган.
У всех млекопитающих имеется вторичное небо; поэтому носовая полость оказывается разобщенной с полостью рта и открывается далеко кзади под основанием черепа в глотку через вторичные хоаны. В качестве рудимента первичной хоаны можно рассматривать существующий у ряда млекопитающих ductus nasopalatinus.
|
The requested URL /down.htm was not found on this server.